Управление большими системами
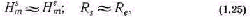
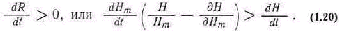
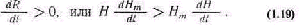
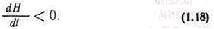
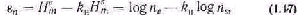
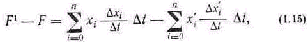
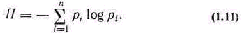
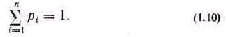
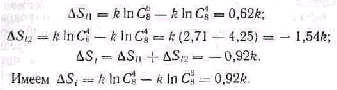

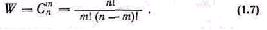

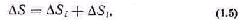

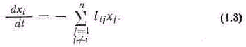
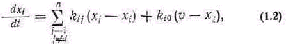
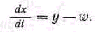
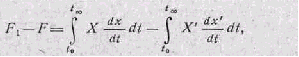
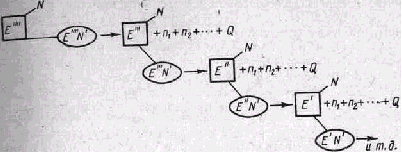
 Управление большими системами
Управление большими системами
Теория автоматического управления разрабатывалась главным образом применительно к производственным процессам и техниче ским устройствам, о которых известны принципы их организации, структура составляющих устройств элементов, порядок связей меж ду ними, закономерности взаимодействия и основные параметры. Такие системы легко поддаются строгому математическому описа нию, их можно однозначно формализовать и управлять на основе определенных алгоритмов. Однако современная техника автомати ческого управления все чаще сталкивается с задачами управления большими системами, к которым ввиду их чрезвычайной сложности такой подход оказывается неприменимым.
Рис. 11. Взаимосвязи между компа ниями, входящими в Северо-восточ ную энергосистему (по А. Коуту, 1970):
/—«Онтарио Хайдро», 2—«Найэгара Моухок», 3 — «Пенсильвания-Нью-Джер-си Мэриленд», 4 — «Консолидэйтед Эди сон К°», 5 — «Конвекс», 6 — «Лонг Айленд Лайтинг», 7 — «Нью-Инглед электрик си стем», 8 — «Нью-Йорк гэс энд электрик корпорейшн», 9 — «Сентрал Хадсон гэс энд электрик корпореишн», 10 — «Орэндж энд Роклэнд ютилити инкорпорейтед», 11— «Бостон Эдисон», 12—«Нью Бэлфорд гэс энд электрик», 13 — «Монтеан электрик», 14 — «Дейтронт Эдисон», 15 — «Консамерс пауэр», 16 — «Хайдро Квебек», 17—«Сент рал Мейн пауэр»
В результате сложные большие системы часто оказываются не управляемыми и в них могут возникать непредвиденные «стихийные» изменения, порой принимающие катастрофический характер.
Ярким примером такой катастрофы может служить выход из строя грандиозной североамериканской энергосистемы Кэнюз (от первых букв слова Canada, Unated States). Эта чрезвычайно боль шая система образовалась на основе соглашений между десятками частных корпораций, фирм и компаний с участием государствен ных организаций. О сложности ее внутренних связей дает пред ставление рис. 11. Расстройство деятельности такого «невидимого робота» (А. Коут, 1970) привело к тому, что с вечера 9 ноября 1965 г. до утра 10 ноября десятки миллионов жителей на громадной территории США и Канады, включая такой гигантский город, как Нью-Йорк, оказались в темноте с парализованным энергохозяйст вом. Внезапно остановилось метро с 600000 пассажиров, которые стали выбираться через тоннели. Однако в полночь еще 10000 из них не могли выйти из вагонов, а 700 оказались в поезде, остано вившемся над рекой. Застряли между этажами лифты в небоскре бах, прекратилось железнодорожное движение, теле- и радиопере дачи, подача газа, работа водопровода, прервалась телефонная и телеграфная связь и т. д. Все началось с того, что одна перегру женная северная линия была выключена защитным реле.Электроэнергия устремляясь по другим линиям, работавшим на пределе, выключила и их одну за другой. В результате подскочила нагрузка на линии, связывающей северную группу с электростанциями дру гих компаний, что вызвало последовательный разрыв соединений между ними. Прекращение взаимной компенсации потребле ния энергии, резкие колебания нагрузки и нарушение синхрон ности в работе генераторов разных электростанций привело к ряду крупных аварий, которые в конечном итоге вывели из строя всю систему энергоснабжения. Большая система оказалась неуп равляемой.
Особенность больших систем состоит не только в очень боль шом количестве элементов, но и во множестве разнообразных свя зей между ними, образующих иерархию подсистем, структура которых может изменяться в зависимости от многих условий. По этому управление такой системой на основе строгого математиче ского описания практически невозможно, так как все эти условия нельзя предусмотреть, а если даже удалось бы их принять во вни мание, то объем необходимых вычислений оказался бы непомер ным, требующим работы самых быстродействующих ЭЦВМ в тече ние многих лет для выдачи командного сигнала. Поэтому управ ление большими системами в отличие от управления обычными, допускающими поэлементное математическое описание строится на основе специальных методов теории операций и сетевого модели рования, теории массового обслуживания и статистического моде лирования.
Теория операций исследует принципы оптимального управления деятельностью коллективов, составляющих большую систему, ко торая стремится к достижению определенного результата. Приме рами операций больших систем может служить борьба с эпиде мией, наступление на фронте, строительство нового города и т. д. Оптимальность управления операцией оценивается по критерию ее эффективности, определение которого составляет важную, но не всегда ясную задачу исследования. Так, в приведенных примерах не вызывает сомнений, что эффективность противоэпидемических мероприятий выражается прекращением заболеваний, а эффектив ность сражения с войсками противника—победой. Однако критерии эффективности градостроения могут быть разные в зависимости от задач обеспечения жилплощадью определенных контингентов на селения или оптимального расположения жилых массивов относи тельно производственных комплексов или максимального исполь зования ограниченных участков застройки и т. д.
Оптимальный план управления операцией вырабатывается пу тем анализа ее математической модели. Такие модели имеют ха рактер сетей, в которых отдельные компоненты операции изобра жаются в виде связок сети. Сетевое моделирование представляет операцию в форме ориентированного конечного графа. Каждое ребро графа представляет собой процессы, позволяющие перейти от одного этапа операции как события, обозначенного соответст вующей вершиной графа, к следующему. Если такой сетевой график организует только сроки проведения операции, то- каждое реб ро графа получает временную характеристику, если, кроме того, на учет берется рациональное использование материальных ресур сов и другие показатели, то вводятся дополнительные характери стики.
Сетевые модели могут иметь фиксированную каноническую структуру, когда во всех вершинах графа выполняется логическая операция «и», т. е. для осуществления события необходимо завер шение всех обусловливающих процессов. Возможна и переменная (альтернативная) структура сети, когда на некоторых вершинах графа выполняется логическая операция «или», т. е. событие может
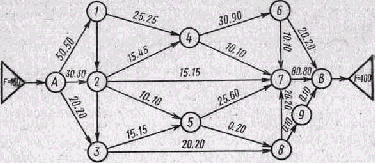
Рис. 12. Сетевая модель для решения транспортной задачи оптимального рас пределения грузопотоков в сложной системе путей (пояснения в тексте)
осуществиться при завершении лишь одного из нескольких обуслов ливающих процессов. Все параметры сети могут существенно изме няться при функционировании большой системы в режиме оператив ного управления, когда на основании сравнения планируемого хода операции с ее фактическим состоянием производится коррекция;
управляющих воздействий.
Типичным примером сетевого моделирования с использованием:
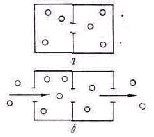
графов может служить транспортная задача оптимального распре деления грузопотоков в сложной системе путей (железнодорожных, автомобильных, авиационных) с учетом их пропускной способности. На рис. 12 показан случай, когда из места отправления (Л) в мес то назначения (В) можно посылать грузы в разных количествах по разным направлениям через различные промежуточные пункты, •в которых происходит пересечение путей. Такая задача решается на основании исходных данных о количестве груза, подлежащего-транспортировке, и пропускной способности каждого участка пути. между промежуточными пунктами 1, 2, 3, 4, 5, 6, 7, 8, 9. Составля ется математическая модель всей операции в виде системы урав нений, в которых выражается процесс максимизации потока гру зов между А и В. По уравнениям этой модели методом линейного программирования находят оптимальное распределение грузопотоков'на участке пути, представленных ребрами графа. Если при нять например, общее количество груза в 100 условных единиц, то при значениях пропускной способности, указанных в виде второй цифры, проставленной на каждом участке пути, доля грузопотока для этого участка при его оптимальном распределении во всей транспортной сети показана в виде первой цифры.
Теория массового обслуживания решает задачи согласования вероятностно-связанной деятельности многочисленных компонен тов большой системы по временным, пространственным, энергетиче ским и другим показателям. Задачи такого рода в технических системах большей частью связаны с фактором времени, в частности с такой организацией средств связи, которая обеспечивала бы удовлетворение вызовов, поступающих в случайном порядке от многочисленных абонентов, или с таким взаимодействием станков разной мощности, при котором пропускная способность всей линии была бы максимальной.
Сложность решения задач массового обслуживания связана со случайным характером взаимодействия элементов больших систем, закономерности которого не поддаются аналитической формализа ции. Поэтому здесь находят применение методы статистического моделирования, в частности так называемый метод Монте-Карло. Используя принципы теории вероятностей, метод Монте-Карло со стоит в воспроизведении большого количества реализаций исследу емого случайного процесса для получения численных значений его искомых характеристик.
Многие особенности управления в больших системах связаны с их иерархической структурой. Такая структура складывается в природных системах, как естественный результат расчленения мно жества взаимодействующих элементов на локальные группы, фор мирующиеся различием местных условий. Каждая такая группа приобретает некоторые специфические черты своей организации и свойств регулирования происходящих в ней процессов. Так форми руются, например, климатические зоны или геологические регионы. Как части большой системы планетарного регулирования они под чиняются его общим закономерностям, но в пределах своего уровня обладают известной степенью автономности.
Иерархическая структура позволяет разделить грандиозную и практически невыполнимую задачу непосредственного управления всеми уровнями подсистем и множествами элементов большой си стемы из единого центра на последовательные операции задания Целевых функций от уровня к уровню. Вместо того чтобы директор крупного завода давал задание каждому рабочему каждого цеха каждый день, он лишь утверждает планы работы цехов, начальник Цеха организует работу бригад, а бригадир управляет деятель ностью членов бригады. При этом решения на каждом уровне при нимаются в результате обработки такого объема информации, ко торый вполне доступен принимающему решение, так как на Уровень более высокого ранга управления передается лишь отфильт-- рованная от ненужных деталей, обобщенная информация. Устрой ства автоматического управления большими системами, построен ные по иерархическому принципу, выгодно отличаются от устройств прямого централизованного управления реальностью и эффектив ностью решения сложных управленческих задач. Такое управление более пластично, так как может гибко изменяться на многих уров нях и более надежно, так как ошибки на одном из уровней могут быть исправлены на других уровнях.
Структурные и функциональные особенности организации биологических систем
Определение живых систем как сложных и очень сложных веро ятностных кибернетических дает основание для детального анализа их структурной и функциональной организации специальными ме тодами кибернетики.
Самоорганизация и ее структурные основания
Прежде всего следует рассмотреть фундаментальное свойство живого—способность к самоорганизации. Противоречия в опреде лении самоорганизации получили образное выражение в следующем высказывании крупного кибернетика У. Эшби (1966) на специаль ном симпозиуме по самоорганизующимся системам: «Так как ни об одной системе нельзя утверждать, что она является самоорганизу ющей и так как выражение «самоорганизующаяся» ведет к укоре нению весьма путаного противоречивого представления о данной проблеме, это выражение, вероятно, вообще не следовало бы упо треблять». Однако на том же симпозиуме Эшби заявил, что «в на стоящее время принципы, лежащие в основе самоорганизующихся систем, известны достаточно полно в том смысле, что над большей частью вопросов приподнята завеса таинственности». Понятие са моорганизации охватывает в наиболее общем виде все специфиче ские свойства жизни—сохранение индивидуальности при непрерыв ном обмене веществ и энергии с окружающей средой, активация с восстановлением исходного состояния при раздражении, воспроиз ведение себе подобных при размножении и т. д.
Самоорганизация характерна именно для сложных и очень сложных вероятностных систем. Структурным основанием самоор ганизации является множественность элементов и разветвленность связей между ними, ведущих к возникновению целостности, а функ циональным основанием — развитие гибкого взаимодействия между элементами по типу обратных связей, направленных на оптимиза цию системы. Зачатки самоорганизации можно встретить и в слож ных вероятностных системах неживой природы. Например, множе ство молекул соли, случайно взаимодействующих в растворе, при достижении определенных условий самоорганизуются в кристаллическое тело. Однако вряд ли нужно перечислять различия между ростом кристалла и ростом живого организма. На уровне живого самоорганизация приобретает важную качественную особенность — она становится способом существования этого класса систем. По этому некоторые, наиболее общие характерные черты биологиче ской самоорганизации проявляются уже в процессе возникновения жизни и связаны с проблемой ее происхождения.
В настоящее время наиболее обоснованной гипотезой о проис хождении жизни является представление А. И. Опарина (1957) о первичной агрегации органических полимеров в коацерватные капли, которые способны к избирательному поглощению опреде ленных соединений из внешней среды, т. е. к зачаточному обмену веществ. Такие коацерватные капли образуются, когда органиче ские молекулы достигают в процессе полимеризации определенных. размеров. Тогда они входят между собой в особые физико-хими ческие отношения, которые дают им возможность выделиться из общего водного раствора.
Обособление системы от окружающей среды составляет сущест венный признак самоорганизации. В современной цитологии все бо лее распространяется мнение, что комплексные коацерваты состав ляют основу протоплазмы живых клеток (А. С. Трошин, 1956). Это находит подтверждение и в том, что, применяя физические и хими ческие воздействия, вызывающие вакуолизацию живых клеток, можно было вызвать явления «вакуолизации» в комплексных коа-церватахФункциональные основы самоорганизации
Казалось бы, что обособление от окружающей среды должно приводить к изоляции системы. Однако в данном случае этого не происходит. Наоборот, выделившаяся из однородной среды самоор ганизующаяся система начинает с ней активно взаимодействовать. Это обусловлено функциональными особенностями образовавшейся таким путем биокибернетической системы связей.
Взаимодействие коацерватной капли с окружающим раствором вначале имеет характер преимущественного извлечения и концент-рирования в ней высокополимерных соединений. Однако в дальней шем из множества полимеров внутри капли возникают сложные вторичные структуры, между которыми также происходит физико-химическое взаимодействие. Внутрисистемные процессы связыва ются с отношениями коацервата и среды и обусловливают непре рывный поток веществ через него — прообраз биологического обме на веществ.
Вся эта сложная эволюция протобионтов наглядно демонстри рует функциональные особенности их химизма, выражающиеся в способности к активному обмену веществ. По-видимому, коацерват-ный агрегат возникает из случайного «зацепления» нескольких мак ромолекул. Однако его внешние и внутренние связи таковы, что однажды возникнув, он вовлекает в свою структуру все больше мак ромолекул, увеличиваясь в размерах и усложняя свою организа цию до какого-то оптимального предела, превышение которого включает тормозные механизмы. Сложившаяся при этом струк турная организация определяет направление и объем проходящего через нее потока веществ, который в свою очередь может влиять на структуру через их пластическое обеспечение. При колоссальном разнообразии структурной организации и вещественного состава коацерватных комплексов вероятность возникновения и развития их прогрессивных форм достаточно велика для действия естествен ного отбора как фактора первичной эволюции.
Обратные связи в живых системах
Важным основанием биологической самоорганизации является обусловленное чрезвычайной разветвленностью структуры исклю чительное богатство и разнообразие обратных связей на всех уров нях живых систем.
Отрицательные обратные связи обеспечивают стабильность функций организма, постоянство его параметров, устойчивость к внешним воздействиям. Они являются основным механизмом гомео-стаза, энергетического и метаболического баланса, контроля чис ленности популяций, саморегуляции эволюционного процесса.
Положительные обратные связи играют позитивную роль уси лителей процессов жизнедеятельности. Особенное значение они имеют для роста и развития. Чем больше живая масса организма, тем больше его ассимиляторные возможности. Примером положи тельной обратной связи в организме может служить также гумо ральная саморегуляция желудочного сокоотделения, когда всасы вание продуктов переваривания белков, возбуждая железы, про грессивно увеличивает переваривание. Вместе с тем положитель ные обратные связи нередко выступают как механизм так называе мого «порочного круга», когда болезнетворные воздействия, нару шающие норму, вызывают в организме изменения, еще более благо приятствующие их действию. Например, сердечная недостаточность ухудшает кровоснабжение миокарда и еще более ослабляет его сокращения. Если отрицательная обратная связь способствует вос становлению исходного состояния, то положительная обратная связь уводит организм и его функции все дальше от исходного состояния.
Взаимодействие положительных и отрицательных обратных свя зей ярко выступает на примере формирования растительных и жи вотных ценозов. С увеличением их биомассы развитие ценоза уси ливается. Однако это усиление имеет место лишь до известных пределов, когда вступает в действие ограничительный механизм саморегуляции и положительная обратная связь сменяется отрица тельной. Можно предположить, что биологическая самоорганизация на всех уровнях—метаболическом, клеточном, тканевом, органном, организменном и видовом—начинается на основе механизма поло жительной обратной связи, на которые затем накладываются огра ничения регуляторных отрицательных обратных связей (см. схему).
Взаимодействие положительных (+) и отрицательных (—) обратных связей в системе регуляции динамики численности популяции
Обратная связь приобретает особое значение именно для си стем биологического типа потому, что такого рода регулятор авто матически компенсирует любые возмущения, даже такие, природа которых неизвестна. Если в простых системах, структура которых может быть точно описана и поведение однозначно предсказано, возможны и другие способы управления путем одностороннего воз действия на отдельные их элементы и звенья, то в очень сложной вероятностной системе, не поддающейся детальному описанию, это единственный способ эффективного регулирования.
По образному определению Н. Винера (1958), обратные связи в живом организме обеспечивают его способность регулировать будущее поведение прошлым выполнением приказов.
Устойчивое термодинамическое неравновесие
Характерной отличительной особенностью живой организации является ее термодинамическое своеобразие. Давно известный зна менательный факт—обязательное наличие основного обмена ве ществ у любого живого существа—убедительно свидетельствует о том, что жизнь должна непрерывно поддерживаться тратой энер гии. По этому поводу Хилл (Hill, 1929) писал: «Живая клетка представляет собой сложную организованную систему..., которая в термодинамическом отношении бесконечно невероятна и может находиться в этом состоянии лишь до тех пор, пока может быть использована свободная энергия для поддержания этой организа ции». Тем не менее в биологии долгое время господствовало и дает о себе знать до сих пор представление о внутренней уравновешен ности живой системы. Однако обобщение обширного фактического материала и развитие теоретической биологии привело к форму лировке Э. С. Бауэром (1935) принципа устойчивого неравновесия как основного принципа термодинамической организации живого. При этом живое состояние характеризуется тремя условиями:
наличие свободной энергии, разряжающейся без внешнего воз действия, т. е. свойства «спонтанной» деятельности; 2) ответ на • внешние воздействия, выравнивающие градиент энергии, восстанов лением этого градиента, т. е. свойства раздражимости и возбуди мости; 3) накопление свободной энергии путем работы против факторов, ведущих к равновесию, т. е. свойства целесообразного поведения и приспособительной изменчивости. Э. С Бауэр рассматривает принцип устойчивого неравновесия как всеобщий закон существования биологических систем и опре деляет его следующим образом: «Все и только живые системы ни когда не бывают в равновесии и исполняют за счет своей свободной энергии постоянно работу против равновесия, требуемого законами физики и химии при существующих внешних условиях». Так, орга низм использует химическую энергию питательных веществ для создания и поддержания внутреннего структурного неравновесия как источника свободной энергии жизнедеятельности, направлен ной на оптимизацию условий существования, в частности увеличе
Рис. 13. Схема метаболических циклов поддержания устойчивого состояния жи вой материи (по Э. С. Бауэру, 1935):
Е — молекулы живой материи (число черточек обозначает степень деформированности ак тивной молекулы), N—молекула питательного вещества. п1, пг—продукты распада, О— выделение тепла
ния притока питательных веществ и, следовательно, на достижение еще большего неравновесия с окружающей средой (рис. 13).
Живая система способна удаляться от равновесного состояния в большей степени, чем любая неживая в равных условиях. Поэтому свободная энергия неживой системы будет больше, чем таковая у энергетически эквивалентной живой системы после того, как они обе выполнят максимальную работу. Разница энергетических уров ней может служить количественной оценкой работы живой системы против равновесия:
где свободная энергия неживой системы обозначается как F1, а жи вой—как F; исходная энергия неживой системы X', а живой—X;
изменения за время t энергетического уровня неживой системы х', Q Живой — х.
Неравновесное состояние, характерное для живой системы, ко-Ренным образом отличается от динамического равновесия, т. е. непрерывного балансирования прихода и расхода, которое часто считают ведущим принципом живой организации. В качестве наи более яркого проявления роли динамического равновесия в жизни животного обычно приводят весьма непродолжительные сроки пол ного вещественного обновления его тканей и органов при внешней неизменности их морфологии и бесперебойном выполнении и\ функций.
Однако такое динамическое равновесие можно наблюдать и в неживой природе. Например, бьющий из-под земли фонтанчик род никовой воды сохраняет свою форму и непрерывно выполняет одну и ту же физическую работу, хотя каждое мгновение его образуют все новые и новые частицы воды, сменяющие друг друга. Особен ность живой системы состоит в том, что она способна использовать внешние ресурсы для создания внутреннего источника свободной энергии, направляемой на работу против внешних физических и химических факторов, а неживая система такой способностью не обладает. Поэтому голодающее животное развивает бурную деятельность по добыванию пищи, а лишенный напора родниковый фонтанчик немедленно сходит на нет.
Негэнтропия против энтропии
Термодинамический аспект изучения биокибернетической систе мы позволил ввести ряд специальных количественных характери стик ее организации. Так, по способности различных видов энергии к превращениям в другие формы Л. Бриллюэн (1960) оценил наи более высоко механическую и электрическую энергию, на сред нее место поставил химическую и на последнее—тепловую. При нимая в качестве меры неупорядоченности тепловой энергии энтро пию (5), а для оценки степени упорядоченности—негэнтропию (N), он предложил количественную характеристику организации си стемы исходя из того, что высококачественные виды энергии отра жают удельный вес упорядоченных, а низкокачественные — случай ных отношений между элементами системы.
Открытая неравновесная живая система находится под совокуп ным влиянием термодинамических, физических и химических зако нов, которые по-разному изменяют величины N и S. Так, второй закон термодинамики допускает лишь такие преобразования энер гии, которые ведут к необратимому накоплению тепловой, т. е. спо собствуют увеличению энтропии, а следовательно, снижают степень организованности системы.
Использование различных физических превращений, особенно при высокой связности элементов системы, может лишь задержать рост энтропии и уменьшение негэнтропии, но не в состоянии оста новить дезорганизацию системы. И только многостадийные хими ческие процессы основного обмена веществ, которые могут проте кать как по эндотермическому (с поглощением тепла), так и по эк зотермическому (с выделением тепла) типу и изменять структуру физических связей, способны использовать низкокачественную теп ловую энергию для производства высококачественной ее формы, т. е. уменьшать энтропию, увеличивать негэнтропию, а следовательно:, повышать организованность системы за счет углубления энтропии
в окружающей среде.
С этой точки зрения становятся понятными условия возникнове ния свойств самоорганизации в сложных вероятностных биокибер-
нетических системах. Сложность и многообразие химических реак ций обусловливают преобразование потоков тепловой энергии в хи мические и физические виды энергии упорядоченных связей При этом наступает повышение химической и физической связности, что ведет к дальнейшему уменьшению энтропии и тем самым еще более усиливает процессы упорядоченности структур системы На этом этапе действует механизм поло жительной обратной связи. Одна ко по мере возрастания организо ванности системы в ней сокраща ется объем неупорядоченно миг рирующей энергии. Эти отноше ния по типу отрицательной об ратной связи удерживают пока затель организации от превыше ния оптимального уровня.
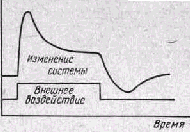
Рис 14 Реакция открытой систе мы на внешнее воздействие (по Burton, 1955)
С открытым характером жи вых систем связано и своеобраз ное течение их реакций на внеш ние воздействия. Если в закры тых системах изменение, например, активности ферментов, дейст вующих на различные звенья цепи химических превращений, мо жет лишь замедлить или ускорить достижение определенного ста ционарного состояния, то в открытой системе такое изменение не только повлияет на скорость реакции, но и приведет систему к но вому положению «равновесия» Причем это новое стационарное положение будет достигаться не прямым путем через промежуточ ные значения, а путем предварительного «выброса» в экстремаль ное состояние (рис. 14). Однако, как указывает И. И. Шмальгау-зен (1961), «все биологические системы являются ограниченно ог-крытыми», так как каналы их связи с внешней средой контролиру ются и регулируются самой системой в соответствии с ее состоя нием. Это действительно для всех уровней биологической органи зации. Так, обмен веществ клетки во многом зависит от влияний нуклеопротеидов ядра и активности энзимов, поведение особи жи вотного от состояния его нервной системы, голода, насыщения, половых мотиваций, взаимодействие вида с окружающей средой — от внутривидового естественного отбора.
Иерархическая организация
Существенной особенностью организации биокибернетических сиcтeм является их иерархическое строение. Как уже было сказано выше, любая система одновременно является элементом системы более высокого уровня, а любой элемент может также рассматриваться как система из более простых элементов. Например, хлоро пласт—это элемент системы растительной клетки, сама клетка—. элемент системы листа, стебля или другого органа растения, в свою очередь лист—элемент растительного организма, а растение в це лом—элемент системы биоценоза.
Каждая система более высокого уровня объединяет в единое целое, прямо или косвенно подчиняет своим закономерностям все нижележащие. Верхние и нижние границы такого иерархического ряда биологической организации определяются переходом к иным категориям форм движения материи. Например, если химический элемент системы обмена веществ в клетке в свою очередь предста вить как систему, организованную из атомов, то это уже не биоло гическая, а химическая система. С другой стороны, системы жизни, связанные с деятельностью человека, выходят из рамок чисто био логических, и на них накладывают свой отпечаток социальные зако номерности.
Управление в биокибернетических системах как целесообразное саморегулирование
Сложная динамическая организация биокибернетической систе мы требует непрерывного управления, без которого система не мо жет существовать. Особенность этого управления состоит в том, что оно происходит на основе самоорганизации и приобретает активный характер.
Активность живых систем
Возможность поддержания определенного состояния в условиях варьирования многих переменных в саморегулирующейся на основе обратных связей системе наглядно демонстрирует гомеостат (У. Эшби, 1962). В живых организмах способность противостоять внешним воздействиям достигает степени активного поведения.
Для живых систем, как мы уже видели выше, особенно харак терны процессы саморегуляции за счет непрерывной внутренней работы создания негэнтропии и неравновесных структур. При этом система не только противодействует влиянию факторов, ведущих к ее дезорганизации, и облегчает действие факторов, благоприят ствующих повышению ее организованности, но в отсутствие тех и других факторов, движимая громадным объемом внутренней орга низующей деятельности, может проявлять независимую от внешних условий, в данный момент обусловленную внутренними факторами так называемую спонтанную активность. Закрепление спонтанной активности во вновь возникающих структурах дает основу явлени ям развития.
Это исключительное свойство живого долгое время служило поводом для виталистических рассуждений и лишь в свете концеп ции современной теоретической биологии и физиологии находит свое место среди естественнонаучных понятий. Исключительнаяструктурная сложность и множественность обратных связей в био-кибернетических системах превращает процессы управления ими в процессы внутреннего саморегулирования по собственным законам организации системы, а термодинамическая неравновесность ведет к их проявлениям в форме спонтанной активности развития.
Поскольку процессы управления в живых системах осуществля ются главным образом как внутренние саморегуляторные, то они определяются в основном внутренними законами биокибернетиче ской организации. Эти законы, как было выяснено выше, обуслов ливают преимущественное направление процессов преобразования системы в сторону возрастания упорядоченности и связаности ее элементов в структурном смысле и снижения значения энтропии в термодинамическом смысле. А. И. Берг (1963) считает даже, что все акты управления в кибернетических системах сопровождаются уменьшением энтропии.
Целесообразность саморегуляции
Из изложенного выше следует, что саморегулирование живой природы осуществляется не хаотично, а в определенном направле нии, в виде решения задачи оптимизации ее биокибернетических систем. Большая сложность и вероятностное строение последних позволяют ей решать задачу оптимизации разными способами, а контроль решения через обратные связи стимулирует поиски но вых способов до получения результата. С этой точки зрения получа ет биокибернетическое определение такое специфическое понятие биологии, как целесообразность, которому также долгое время при давали виталистический оттенок. По-видимому, в понятиях термо динамики целью живой системы следует считать оптимизацию ее поведения в данных условиях, которая достигается путем увеличе ния структурной и энергетической неравновесности со средой, вы ражаемой функцией отношения негэнтропии к энтропии. Однако ввиду сложности биологической эволюции не всегда можно точно определить критерии такой оптимизации. Целесообразная саморе гуляция является основной формой процессов управления в живой природе.
Цели системы биосферы и ее подсистем
Представление о целесообразности саморегулирования в живых системах, как механизме их приспособления к конкретным услови ям существования освобождает понятие цели от телеологических и антропоморфических толкований. Такой взгляд находит поддерж ку и среди философов (В. С. Украинцев, 1973). Исходя из этого представления цель биологической системы можно определить, как объективно проявляющееся направление активности, зависящее от ее организации и влияний окружающей среды.
Как уже было отмечено, общее направление активности биосфе ры в целом идет в сторону повышения уровня организации и накоп ления свободной энергии устойчивого неравновесия. Однако роль основных подсистем земной жизни в достижении этой цели оказы вается весьма различной.
Зеленые растения, используя силы (солнечная энергия) и мате риалы (углекислота, вода, соли) неживой природы, создают пер вичные органические соединения, несущие энергию устойчивого неравновесия. Они создали и продолжают поддерживать атмосфе ру нашей планеты и являются базой для существования более вы соких форм жизни. Поэтому направление активности или цель растительной подсистемы биосферы можно определить как первич ный синтез биомассы из неорганических источников и создание ис ходного негэнтропийного материала.
Масса живого вещества, созданная растениями, используется животными прямо (травоядные) или вторично (плотоядные) для преобразований в более высокооргаяизованные структуры свое го тела. На основе этих преобразований возникают такие новые вы сокоэффективные приспособительные функции, как двигательная и нервная, резко увеличивающие активность организмов. Общее
направление активности в животной подсистеме биосферы можно определить как прогрессивные преобразования биомассы, повыша ющие ее структурную организацию и уровень негэнтропии.
Принципиальная особенность человеческой деятельности состо ит, как известно, в использовании орудий труда. Тем самым впер вые в истории развития жизни на Земле создание негэнтропии и преобразование биомассы было вынесено за пределы живого орга низма и стало совершаться в искусственно организованных произ водственных процессах, использующими материалы и силы как живой, так и неживой природы. Если растения и животные огра ничены в использовании природных ресурсов «пропускной способ ностью» обмена веществ в своих организмах, то человек снимает для себя эти ограничения и «всю природу превращает в его неорга ническое тело» (К. Маркс, 1844) *. В отличие от растительной и жи вотной цель человеческой подсистемы биосферы можно определить, как создание посредством орудий труда позволяющих создавать небиологическим техническим путем свободную энергию негэнтро пии в искусственных системах, воспроизводящих процессы, осуще-
ствляемые до того лишь живой материей. Растения и животные вы рабатывают термодинамическую и структурную энтропию в себе,
•человек вырабатывает ее машинами для себя.
Классификация механизмов саморегуляции
Классификация видов саморегулирования в живых системах
еще мало разработана. С. Н. Брайнес и В. Б. Свечинский (1963) предложили общую схему управления функциями организма на трех уровнях. Низший уровень нервной и гуморальной регуляции
«обеспечивает по интероцептивным сигналам постоянство основных
* См.: Маркс К., Энгельс Ф. Из ранних произведений. М., Госполит-/издат, 1956, с. 565.
(физиологнческих констант (кровяного давления, состава крови, температуры тела и т. д.). Средний уровень безусловнорефлектор-ного управления обеспечивает приспособительные реакции организ ма в связи с изменениями его состояния (например, поведение голодного и сытого животного). Высший уровень условнорефлектор-ного управления обеспечивает по сигналам внешнего мира приспо собительные изменения вегетативной сферы и целостного поведения организма.
Анализ процессов управления с учетом термодинамических кри териев привел к выделению семи уровней целесообразного саморе гулирования в ответ на- внешние так называемые энтропийные-факторы, повышающие внутреннюю энтропию живой системы-(В. И. Черныш, 1968): 1. Стабилизация путем поддержания посто янными параметров системы при кратковременном действии слабых энтропийных факторов. 2. Адаптация путем перестройки внутрен ней структуры и функций системы при длительном или постоянном действии слабых энтропийных факторов. 3. Самовосстановление путем создания новых структур взамен разрушенных кратковре менным действием сильных энтропийных факторов. 4. Размноже ние путем воспроизведения себе подобных для сохранения системы вида при действии факторов, уничтожающих отдельные особи. 5. Развитие путем объединения и реализации в одной системе всех предыдущих уровней саморегулирования при действии разнообраз ных энтропийных факторов. 6. Эволюция путем переключения на использование новых видов веществ и энергии при действии энтро пийных факторов истощения окружающей среды 7. Соревнование путем формирования структур и развития функций, противодейст вующих влиянию энтропийных факторов, порождаемых деятель ностью других биокибернетических систем.
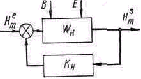
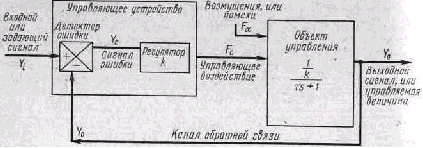
В технике возможно четкое разделение управляющего устройст ва и объекта управления (В. В. Солодовников, 1961). Однако в жи вом организме они настолько переплетаются, что границу между ними бывает трудно провести. Например, когда образующиеся при переваривании пищи пептоны, всасываясь в кровь, вызывают усиленное выделение желудочного сока, который увеличивает обра зование пептонов, то белковые вещества выступают в роли управ ляющего устройства и в роли объекта управления. Более ясное разграничение управляющих и исполнительных структур имеет мес то в системе движений (А. А. Гидиков, 1964). В управляющих воз действиях предыдущих звеньев рефлекторного механизма на по следующие проявляется принцип детерминирующей роли «станции отправления» (Г. Н. Крыжановский, 1975).
Ф. Гродинз (1966) предпринял попытку анализа общих принци пов организации управляющих систем, наиболее выраженных в био логических системах. Он выделил в управляющем устройстве две-подсистемы—детектор ошибки и собственно регулятор. Первая оценивает рассогласование между фактическим выходным и задан ным сигналами. Вторая, получив сигнал ошибки, вычисляет и вы дает на выход скорректированную управляющую команду, которая поступает к управляемому объекту (см. схему). Во всех рефлектор ных механизмах проявляется управляющее воздействие предыду щих звеньев на последующие.
Блок-схема системы управления с обратной связью
(по Ф. Гродинзу)
Некоторые общие свойства биологической саморегуляции
Так как саморегуляция в биокибернетических системах осуще ствляется не каким-либо одним управляющим воздействием, кото рое бы сразу привело управляемый объект в заданное состояние, а состоит из параллельно-последовательных, нередко антагонисти ческих команд, лишь постепенно достигающих цели, процессы уп равления в живых системах, как правило, являются дискретными и периодическими. Поэтому для оценки их свойств важно знать время релаксации, которое определяет соотношение длительности переходных и установившихся режимов работы биологических си стем.
В живой природе эти характеристики оказываются весьма раз личными по величине, но далеко превышающими таковые техниче ских кибернетических систем. Так, переходные режимы рабочих блоков современных быстродействующих электронных вычислитель ных машин измеряются наносекундами, а наиболее короткое время релаксации живого механизма дает рефрактерный период прове дения нервного импульса, составляющий около одной миллисекун ды. Между ними оказывается тысячекратный разрыв. Однако боль шинство биологических систем работает гораздо медленнее нерва. Например, переходные режимы деятельности сердца и сосудов, вызванные депрессорным рефлексом, типичным для гомеостатиче-ских саморегуляций организма, длятся многие минуты, а регуляции на уровне биоценозов типа взаимного контроля численности попу ляций длятся месяцы и годы.
Саморегуляция имеет не только материально-энергетическую, но и информационную сторону и является таким видом движения ин формации, который уменьшает неопределенность системы. Особен ста информационного представления биокибернетических систем определяются прежде всего их большими размерами и богатством вероятностных внутренних связей. Например, чтобы представить количественно способность глаза человека к различению образов, которые могут возникнуть в поле зрения, то даже если упрощенно ппринимать, что каждый из 130 млн. фоторецепторов может нахо диться лишь в двух состояниях, мы получаем число 211.310^8, которое далеко выходит за пределы величин, имеющих реальное значение. Исходя из общего числа атомов в видимой Вселенной примерно 1073 и полагая, что событие на атомном уровне совершается за 10-10 с, У. Эшби (1966) подсчитал максимальное число физических собы тий, которые могли произойти с момента образования земной коры, и получил число 10100. Это наибольшее число, характеризующее ма териальные процессы в нашем мире. Для записи большего числа событий не хватит атомов Вселенной. Отсюда следует нереальность описания каждого из возможных состояний сложной биологической системы для ее регулирования и неизбежность обобщенного стати стического подхода к оценке этих состояний с целью решения задач переработки циркулирующей в них информации и управления эти ми системами. Поэтому процессы нервной деятельности описывают ся в понятиях статистической нейродинамики.
Механизмы эволюции и саморегуляции жизни
Выше было показано, что фундаментальным принципом живого является способность к самоорганизации, направленной на дости жение все более высокой упорядоченности в борьбе с энтропией. В этом направлении и происходит эволюционное развитие мира живых существ.
Биокибернетическое определение эволюции
С точки зрения общих принципов кибернетики эволюция пред ставляет собой процесс прогрессивной оптимизации интегральной системы жизни. Эта оптимизация происходит по указанным выше структурным, функциональным, термодинамическим, информацион ным и другим показателям на всех уровнях иерархической системы жизни — субклеточном, клеточном, органном, индивидуальном, ви довом и биоценотическом. Наступающие на каком-либо из этих Уровней прогрессивные адаптации неизбежно вызывают взаимо обусловленные изменения на всех остальных уровнях. Например, развитие физиологического механизма теплорегуляции обусловило, с одной стороны, глубокие перестройки метаболизма клеток тепло кровных животных, в частности ускорение обменных процессов, а с другой стороны, вызвало коренные изменения в экологии их видов, в частности резкое увеличение ареалов обитания и снятие сезонных ограничений активной деятельности.
Однако не все уровни интегральной биологической системы равноценны по своей роли в механизме эволюции живых существ, происходящих в природе. По мнению И. И. Шмальгаузена (1961) наиболее существенным звеном, от которого как от первичного обычно начинаются эволюционные преобразования, является попу. ляция—элементарная эволюирующая единица. Именно в популя ции впервые возникают перестройки, которые затем распространя ются «вверх» и «вниз» по иерархической структуре жизни.
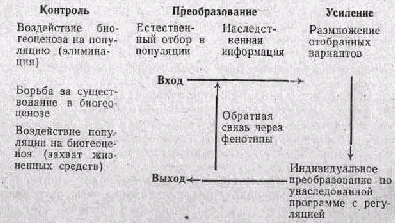
Популяция находится под непрерывным воздействием всей мас сы абиотических и биотических воздействий внешней среды, обо значаемой как биогеоценоз (В. Н. Сукачев, 1945). Как очень слож ная система биогеоценоз также организуется на основе некоторых •общих принципов взаимодействия его элементов, из которых важ ным фактором эволюции является борьба за существование.
Как видно из приведенной ниже схемы, цикл эволюционных пре образований популяции начинается с воздействия биогеоценоза на популяцию путем прямого и косвенного истребления ее особей. Этот входной канал несет информацию о состоянии внешней среды. Затем происходит естественный отбор фенотипов внутри популяции и меняется наследственная структура. Путем размножения новые свойства усиливаются и генетически закрепляются. Наследственная информация реализуется в новой фенотипической форме. Наконец, преобразованная популяция образует выходной канал, несущий информацию о ее состоянии путем активной деятельности особей, захватывающих жизненные средства из биогеоценоза.
Кибернетическая схема регуляции эволюционного процесса
(по И. И. Шмальгаузену, 1961)
В зависимости от приспособительного значения наступивших из менений обратная связь через фенотип может иметь разное на правление. Более приспособленные фенотипы размножаются, и эта движущая форма естественного отбора является механизмом положительной обратной связи. В случаях, когда новые фенотипы оказываются менее приспособленными, они гибнут, происходит воз вращение к прежнему фенотипу, и такая стабилизирующая форма естественного отбора является механизмом отрицательной обрат ной связи.
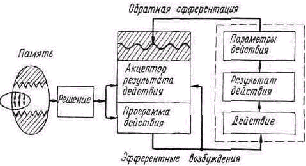
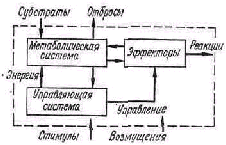
Открытый характер живых систем. Для понимания процессов, протекающих в биосистеме, необходимо учи тывать две стороны ее функционирования. Одна из них связана с открытым характером системы — это процессы получения, накопления, передачи и использования ве ществ, энергии и информации. Эти процессы обеспечива ют возможность сохранения структуры, рост и выполне ние всех специфических функций биологической сис темы.
Рис. 1. Схема потоков энергии и информации в организме (по Т. Уотермену).
Другая сторона функционирования, связанная с уп равлением, включает восприятие, хранение, переработку и использование информации. Информационно-управля ющие механизмы в системе определя ют, какие вещественные и энергети ческие процессы и с какой скоростью происходят в ней. Наиболее общей за дачей управляющих систем организма является сохране ние его основы, со здание благоприят ных условий для ее
функционирования при изменяющихся условиях внеш ней среды. Как отмечает Г. Уотермен, кибернетические механизмы для того и существуют, чтобы обеспечить ста билизацию и сохранение энергетической части организ ма [132].
На рис. 1 показана схема потоков энергии и информа ции в организме животного. Энергетическая часть орга низма обозначена как метаболическая система, а управля ющая часть содержит три блока — генетическое управле ние, физиологическое управление, эффекторы.
Структура организма поддерживается механизмами генетического управления. Получая от остальных систем энергию и информацию, генетическая система управляет процессами синтеза необходимых веществ и поддержива ет жизнедеятельность остальных систем организма. Процессы в генетической части протекают достаточно медленно, для нее характерны долгие интервалы времени, связанные с процессами роста, становления организма и его старения, процессы регенерации тканей и другие. Поведенческие реакции организма осуществляются си стемой физиологического управления. Процессы в физи ологических системах управления организма протекают значительно быстрее, чем в генетической. Поэтому гене тическая система образует структуру биосистемы, в то время как быстрые поведенческие и физиологические процессы определяют ее функцию. Организация живого есть единство структуры и функции — устойчивости и подвижности. «То, что называется структурой, является медленным процессом большой продолжительности: то, что называют функцией, является быстрыми процессами короткой продолжительности» [121]. Таким образом, само строение биосистемы отражает две главнейшие ее характеристики — процессы обмена веществ (открытый характер живых систем) и процессы управления.
Понятие открытой системы впервые было введено в обиход биологической науки Л. фон Берталанфи в 1932 г. [38]. Для такой системы характерно, что в нее постоянно извне вводятся вещества, которые внутри системы под вергаются различным преобразованиям. В результате процессов синтеза (анаболизма) в системе возникают компоненты более высокой сложности, утилизируемые организмом. Одновременно происходят процессы распада (катаболизма), конечные продукты которого выводятся из системы.
Одной из наиболее характерных черт открытых систем является то, что в них достигается состояние под вижного равновесия. При этом структура системы оста ется постоянной, но это постоянство сохраняется в про цессе непрерывного обмена и движения составляющего ее вещества.
Если обозначить общую скорость расходования не которого вещества в биосистеме через w, общую скорость поступления этого вещества через у, а его количество в биосистеме через х, то можно записать очевидное уравне ние
Это уравнение иногда называют уравнением Берта ланфи. В современной литературе [162] открытая систе ма часто представляется в виде совокупности отдельных блоков-компартментов, между которыми происходит об мен веществами. Тогда процессы в открытой системе, вызванные наличием различных концентраций вещест ва в разных компартментах и в окружающей среде, описываются обыкновенными дифференциальными урав нениями. Так, если обмен веществом между компартмен-тами и средой определяется законом диффузии, когда скорость потока пропорциональна разности соответству ющей концентрации вещества, то уравнения системы имеют вид
где п — число компартментов в системе; xi — концентра ция вещества в i-м компартменте; kij — коэффициенты диффузии, i = 1,2,...,п; j = 1, 2, ..., n; kij 0; v — концентрация вещества в окружающей среде; ki0 — ко эффициенты диффузии на границах системы.
Традиционной методологической основой описания биологических систем является термодинамика открытых систем [НО]. Если термодинамическая система не обме нивается со средой веществом, то она называется замкну той.. Изменения динамических переменных х, описыва ющих замкнутую систему вблизи равновесия, подчиня ются так называемым уравнениям Онзагера
Пусть матрица L, составленная из коэффициентов 1ц, имеет только вещественные и отрицательные собственные значения. Это значит, что система (1.3) имеет стационар ное равновесное состояние, которое достигается в ходе апериодического переходного процесса.
Энтропия S в замкнутых системах либо не меняется (при обратимых процессах), либо возрастает при необра тимых
В стационарном состоянии энтропия достигает макси мального значения Smax.
Методы термодинамики открытых систем [110] при меняются для описания систем, обменивающихся веще ством со средой и близких к термодинамическому равно весию. Изменение энтропии в открытой системе включает две компоненты
где Si, Sl — прирост энтропии за счет процессов внут ри системы и приток энтропии извне соответственно. В за висимости от соотношения Si и Sl, величина S может быть положительной и отрицательной. В стационарном состоянии S = 0.
Рис. 2. Компартментальные системы: а— замкнутая; б— открытая.
Если рассматриваемая открытая система близка к тер модинамическому равновесию, то поведение энтропии определяется теоремой Пригожина, согласно которой в стационарном состоянии прирост энтропии, обусловленный проте канием необратимых процессов внутри системы, имеет минималь ное из возможных и положитель ное значение. По этой теореме в уравнении (1.5) в стационарном состоянии величина Si прини мает минимальное, но положи тельное значение.
Для практических задач ме тоды термодинамики открытых систем применяются не часто, бо лее распространены методы ком-партментального анализа. Остановимся на одном простом примере, позволяющем понять связь, существующую между этими двумя подходами.
Рассмотрим сначала замкнутую систему — ящик с двумя отделениями и отверстием в перегородке (рис. 2, а). Такая модель используется иногда для разъяснения ста тистического характера энтропии в замкнутой системе. Обобщим ее на случай открытой системы.
Определим состояние системы как (от, п—т), где п — общее число частиц в ящике, т — число частиц в первом (левом) отсеке.
Энтропия состояния системы
где W — число комбинаций, отвечающих данному состоя нию; k — константа Больцмана. Число комбинаций, отвечающее состоянию (т, п — т) (табл. 2):
В замкнутой системе реализуется стационарное рав новесное состояние (1.3) с максимальной энтропией S = 2,99kl(табл. 2).
Дополним теперь систему двумя внешними потоками частиц: в первый отсек ящика извне поступает поток с за данным темпом — две частицы в единицу времени; из второго отсека — частицы с таким же темпом уходят в окружающую среду. Обозначим потоки символом у с двойным индексом: среде присвоим индекс 0. Тогда y01= 2, y20 = —2.
Пусть исходное состояние системы есть (4,2) (рис. 2, б), а движение частиц между отсеками происходит по закону диффузии
y12 = а [т — (п — m)] = а (2т — п); (1.8) для простоты примем а = 0,5. Рассмотрим теперь дис кретную последовательность событий в системе. После притока двух частиц извне в первый отсек со стояние системы будет (6,2). Затем согласно (1.8) из пер
Таблица 2 Динамика состояний системы
вого отсека во второй перейдут две частицы — система окажется в состоянии (4,4), а после оттока двух частиц из второго отсека в среду — в состоянии (4,2). Переходы (4,2) в (6,2) и (4,4) в (4,2) вызваны обменом со средой, переход (6,2) в (4,4) — внутренними необратимыми про цессами в системе. Вычислим приращение энтропии Si и Sl (1.5).
Приток энтропии извне составляется двумя компонен тами: Sl1 + Sl2, отвечающими переходом системы (4,2) в (6.2) и (4,4) в (4,2). Тогда:
Следовательно, за цикл S = Si + Sl = 0 и си стема находится в стационарном режиме. Начальное и конечное состояния системы совпадают.
Стационарное состояние (4,2) является неравновес ным, оно поддерживается за счет непрерывного протека ния через систему потока частиц. Энтропия стационар ного неравновесного состояния (4,2), равная 2,7lk, меньше максимального значения 2,99k, отвечающего ста ционарному равновесию (3,3).
Кроме этих общих биологических характеристик, це лесообразно рассмотреть качественные состояния, в ко торых может находиться биологическая система.
Норма и патология. Известно, что организм может на ходиться в двух состояниях — нормы и патологии. Эти состояния присущи биосистемам любого иерархического уровня. Состояние нормы является естественным с точки зрения жизнедеятельности. Оно относительно устойчи во и вместе с тем динамично. По отношению к человеку норма выражается в здоровье, и по уставу Всемирной организации здравоохранения определяется как состоя ние полного физического, душевного и социального благо получия, а не только как отсутствие болезней или физи ческих дефектов. Состояние здоровья предусматривает нормальное функционирование биосистем всех иерархи ческих уровней организма. В то же время патология лю бого уровня в силу взаимосвязи и интегрирования всех иерархических систем приводит к патологии всего орга низма. Таким образом: 1) состояние нормы систем всех уровней является необходимым и достаточным условием здоровья всего организма; 2) состояние нормы систем од ного из уровней является необходимым, но недостаточным условием здоровья всего организма; 3) состояние патоло гии систем одного из уровней является необходимым и достаточным условием патологии всего организма. Поопределению И. П. Павлова, патологическое состояние — «это встреча, соприкосновение организма с каким-либо чрезвычайным условием, вернее, с необычным размером ежедневных условий» [101].
Жизнь организма возможна благодаря широкому спектру эволюционно приспособительных реакций, воз никающих в ответ на действие факторов внешней среды. Воздействия окружающей среды на организм, которые могут привести к патологии, условно можно разделить на косвенные и прямые. В результате косвенных влияний среды могут произойти нарушения нормального функци онирования какой-либо системы организма, влекущие за собой снижение резистентности. На этом фоне может развиться патология любой этиологии, в том числе не специфичная для данного фактора среды (например, за болевания, вызванные условно патогенными микроор ганизмами в результате переохлаждения). К прямым воздействиям среды на организм, приводящим к соответ ствующей патологии, можно отнести вещественные (хи мические вещества, микроорганизмы), энергетические (радиация, температура), механические, информацион ные (стрессовые ситуации, иатрогения, самовнушение). Степень отклонения от нормы, вызванная внешними фак торами, зависит от силы и кратности воздействия и от адаптационных возможностей организма. Оно может про являться как в нарушении функции систем любого уров ня иерархии, так и в нарушении структуры составляющих их элементов. Чисто функциональные нарушения явля ются более динамичными и по природе обратимыми. Кум-муляция обратимых функциональных нарушений, свя занная с их продолжительностью и частотой, может при вести и к структурным изменениям, которые более статичны и по природе менее обратимы.
1.2. СВОЙСТВА БИОЛОГИЧЕСКИХ СИСТЕМ
Биологические системы обладают таким обширным на бором свойств, диалектически противоположных друг другу, которых нет у технических и экономических си стем. Это устойчивость и управляемость, жесткость и гибкость, детерминированность и стохастичность и дру гие. В зависимости от обстоятельств у биосистем на пер вый план выдвигаются те свойства, которые необходимыдля сохранения вида и индивида, например обучаемость, изменение поведенческих реакций, приспособление к из менившимся условиям жизни, изменение наследственных качеств. Здесь все подчинено основной цели — выжива нию на уровне организма и воспроизведению на уровне вида.
Одна часть свойств относится к структуре и функции биосистем; другая подчеркивает те качества, которые не обходимы для сохранения вида и индивида; третья ха рактеризует тактику и стратегию организма при достиже нии основных целей.
Сложность и организация. Одним из главных свойств биосистемы является структурная и функциональная сложность. Проявления сложности многообразны и свя заны с количеством возможных состояний. Состояния био системы включают состояния элементов и сочетания свя зей между ними.
У- Р. Эшби [150] предложил в качестве меры слож ности использовать разнообразие, или число состояний системы п. Для оценки сложности системы удобно поль зоваться также логарифмической мерой
Hmax=log n, (1.9)
где Нтах — мера сложности, или максимальная неопре деленность системы.
Функциональные особенности биосистемы проявляются на ее выходе. Поэтому степень функциональной слож ности зависит от характера ее выходных элементов. Если выход биосистемы является дискретной величиной (число выходных значений можно точно пересчитать), то число состояний определяется точно (например, частота нейронов). При этом сложность равна логарифму максимального значения дискретного выхода.
Для биосистем, показатель работы которых изменяет ся непрерывно (например, насыщение крови кислородом, уровень сахара в крови), для получения числа состояний необходимо знать граничные значения изучаемого пока зателя и точность методики его измерения. Таким обра зом, для биосистем, непрерывно изменяющих свой уро вень, число состояний определяется исследователем и является условным.
Оценка сложности по числу состояний системы не по казывает, в каком из возможных состояний система на ходится и в какое она перейдет в следующий момент вре мени. Пусть, находясь в одном из структурных состояний, система принимает любое из п функциональных состоя ний. Если достаточно долго наблюдать систему, то по час тоте появления функциональных состояний можно при ближенно судить о вероятностях ее пребывания в этих состояниях. Обозначим их через pi. Для общей оценки си стемы по вероятностям К. Шеннон ввел эвристическое по нятие неопределенности, или энтропии [145]. Пусть из мерены относительные частоты (при большом числе на блюдений стремящиеся к вероятностям) для всех функ циональных состояний системы. Все функциональные состояния образуют полную систему событий. При этом выполняется соотношение
Под неопределенностью (энтропией) дискретных ве личин понимают следующую величину:
Если, например, система все чаще находится в 1-м состоянии, то вероятность pi стремится к единице. В пре дельном случае, когда pi=1, энтропия равна нулю и система становится детерминированной. Если система безразлична к своим состояниям, то вероятности равны, а неопределенность, вычисленная по формулам типа (1.11), принимает максимальное значение. Таким обра зом, неопределенность системы находится в пределах 0m. (1.12)
Со сложностью связаны многомерность и многосвяз-ность биосистем, проявляющиеся в наличии большого количества разнородных параметров, в многообразии связей между однородными и разнородными параметра ми, характеризующими работу данной системы. Если даже выход биосистемы оценивается по одному параметру, то необходимо учитывать, что этот параметр является результирующим взаимосвязанной работы структур но обособленных подсистем, каждая из которых оцени вается своим параметром. Внутренняя же структура био логических систем организма обусловлена их эволюцией.
С внутренней структурой биологических систем обыч но связывается их организация — специфическая для живых систем структурно-функциональная упорядочен ность. Качественно более сложный уровень организации биосистем по сравнению с естественными системами не органической природы и с искусственными системами, создаваемыми человеком, обусловлен их длительной эволюцией. Формальное определение организации свя зано с работами К. Шеннона, У. Р. Эшби, В. М. Глушко-ва, Г. Ферстера.
Организация системы выявляется путем подсчета меры неопределенности ее состояний, характеризующей ее хаотичность, неорганизованность. При равных вероят ностях принятия системой своих состояний она может счи таться полностью дезорганизованной, так как в любой момент она с равной возможностью может перейти в лю бое состояние. В этом случае система обладает максималь ной неопределенностью, которая совпадает с мерой слож ности системы [1.9]. Можно предположить, что в дезор ганизованной системе ее состояния не связаны между собой.
Оценка уровня организации системы связана с макси мальной Hmax (1.9) и текущей H (1.11) неопределенностя ми системы. Пусть в результате эволюции, фило- или он тогенеза система, работавшая прежде с максимальной неопределенностью Hmax (т. е. полностью дезорганизо ванная система), стала предпочитать некоторые из со стояний и характеризоваться текущей неопределенностью H < Hmax. Тогда организация системы для данного уровня развития определяется реализованной в системе неопределенностью [11,19]:
O=Нmax—Н, (1.13)
где О—абсолютная организация системы. Значение аб солютной организации системы ограничено снизу ну лем, а сверху — величиной, максимально возможной для данной системы неопределенности.
Таким образом, организация детерминированной системы (H=0) также определяется максимальной не определенностью, т. е. строится на максимально возмож ном числе состояний. Только в случае детерминирован ной системы смена состояний является закономерной. Для системы, замкнутой в организационном отношении, равенство (1.13) определяет закон сохранения организа ции: организация и неопределенность на любом этапе эволюции (жизни, обучения и других) равны максимально возможной неопределенности системы. От соотношения (1.13) легко перейти к формуле под счета относительной организации системы R, разделив обе части равенства на Hmax [134]:
R=1-H/Hmax. (1.14)
Очевидно, что мера R характеризует величину отно сительной организации системы и лежит в пределах 0R. Для детерминированной системы относи тельная организация равна единице, для полностью дезорганизованной — нулю. Понятие относительной ор ганизации позволяет сравнивать между собой различ ные системы. Текущее значение неопределенности связа но с энтропией живых систем.
Любая биосистема характеризуется структурной и функциональной организацией. Основой, на которой строится структурная организация биосистемы, являют ся размеры элементов системы, число элементов системы и связей между ними. Например, размеры клеток дан ного нервного узла являются параметрами структурной организации, а по гистограмме распределения клеток данного узла, по их диаметрам можно подсчитать сте пень организации с помощью (1.13) и (1.14). Параметра ми функциональной организации, например отделов нервной системы, могут служить межспайковые интер валы спонтанной и вызванной активности, по гистограммам межспайковых интервалов можно также рассчитать величину абсолютной (1.13) и относительной (1.14) ор ганизации.
Обратимся к понятию информации. Это понятие обыч но предполагает наличие источника и потребителя, объек та и субъекта. Мерой количества информации служит разность неопределенностей, полученная в результате наблюдения объекта, либо разность неопределенностей, получаемая как результат вычитания неопределенности объекта и неопределенности субъективной гипотезы об объекте. Таким образом, как информация, так и органи зация могут выражаться одной и той же мерой и в част ном случае могут иметь одни и те же числовые значения, однако эти значения по-разному интерпретируются. Например, если анализируются материальные объекты без учета взаимодействия между ними, то целесообразно говорить о сложности и организации как свойствах объективных, не зависящих от наблюдателя, исследова теля, как о качествах самих объектов/
Сохранительные свойства. При анализе биосистем обычно сталкиваются с различными аспектами сохрани-тельных свойств живых систем, частными проявлениями которых являются адаптация, выживание, эквифиналь-ность, стабилизация, гомеостаз. Сохраняются только такие составляющие живой природы, в которых стрем ление к самосохранению выражено достаточно ярко [85].
Вообще, термин «сохранительные свойства биосистем» используется для обозначения двух различных, хотя и тесно связанных между собой, характеристик живых систем. Прежде всего самосохранение биосистемы оз начает ее способность поддерживать и сохранять свое стационарное неравновесное состояние вне зависимос ти от условий внешней среды и при изменяющихся условиях функционирования биосистемы. В этом смысле можно утверждать, что управляющие механиз мы биосистемы обеспечивают баланс потоков энтропии через систему, а также баланс вещественных потоков — адекватное снабжение системы необходимыми ей веще ствами и энергией. Не менее важный аспект сохранитель-ных свойств биосистем связан с поддержанием гомео-стаза — относительного постоянства существенных пере менных внутренней среды биосистем при наличии возмущающих влияний со стороны внешней среды.
Управляющие механизмы биосистем, обеспечива ющие их сохранительные свойства, характеризуются высокой надежностью. Надежность сохранительных меха низмов в биосистемах достигается за счет множествен ности (дублирования) регулирующих связей, которые контролируют одну и ту же функцию относительной ав тономности параллельно функционирующих элементов, а также за счет дублирования источников энергии и соз дания запасов нужных веществ.
Одним из важных свойств, позволяющих биосистеме сохранять относительно неизменными параметры ее жиз недеятельности, является гомеостаз, или гомеостазис. Первоначально термин «гомеостаз» был введен У. Кэн-ноном для описания способности организмов поддер живать постоянство внутренней среды: «Постоянные ус ловия, которые поддерживаются в организме, можно было бы назвать равновесием. Это слово, однако, име ет довольно точное значение для относительно простых физико-химических состояний в замкнутых системах, где уравновешиваются известные силы. Координированные физиологические процессы, которые поддержи вают большинство установившихся состояний в организ ме, настолько сложны и специфичны для живых существ, включая, возможно, мозг и нервы, сердце, легкие, почки и печень, работающие совместно, что я предложил спе циальное обозначение для этих состояний — гомеостазис. Это слово не предполагает чего-то установленного и не подвижного, застывшего. Оно означает — состояние, которое может меняться, но которое относительно по стоянно» [154].
Согласно концепции Кэннона, при наличии угрозы со стороны внешней среды в системах организма вклю чаются агенты, уменьшающие эту опасность. Корректи рующие механизмы действуют в основном через специ альную часть нервной системы, функционирующую как регуляторный механизм. Вследствие этой регуляции в организме обеспечивается, во-первых, запас веществ как средство обеспечения соответствия между спросом и предложением, во-вторых, имеются скорости непрерывно протекающих в организме процессов.
Эта концепция была последовательно применена Кэн-ноном для анализа процессов сохранения постоянства содержания воды и солей в крови, поддержания адекват ного кислородного снабжения, гомеостаза сахара, белков, жиров и кальция крови, гомеостаза нейтральности крови "и постоянства температуры тела.
Позже понятие гомеостаза стало формулироваться в более компактной форме: гомеостазом обычно называют сохранение постоянства внутренней среды организма при наличии возмущений во внешней среде.
Отметим также, что термином «гомеостаз» иногда обо значают и саму систему, обладающую гомеостатическими свойствами. Такое использование этого термина восходит к работе Эшби [151]. Приведем в заключение определе ние гомеостаза, данное в стандартном английском меди цинском словаре: «Гомеостаз (1)—состояние равновесия в живом организме, относящееся к различным функци ям и химическому составу жидкостей и тканей, напри мер, к температуре, частоте пульса, давления крови, содержания воды, сахара крови и т. д. (2) — процесс, посредством которого это равновесие поддерживается» [1643.
Идея гомеостаза, предложенная первоначально для физиологических систем, оказалась настолько богатой иплодотворной, что быстро распространилась и на другие биосистемы.
Согласно определению Кэннона, гомеостаз — это свойство целостного организма, целостной биосистемы. Иной точки зрения придерживается Л. фон Берталанфи:
«Следует уяснить, что термин «гомеостаз» может употреб ляться двояко. Он используется либо в его первоначаль ном смысле, предложенном Кэнноном и иллюстрируемом примерами поддержания температуры тела и других физиологических переменных с помощью механизмов об ратной связи, либо в другом смысле, который имеют в ви ду, а именно как синоним для органической регуляции вообще» [38]. Далее Берталанфи настаивает на использо вании термина «гомеостаз» как синонима термина «от рицательная обратная связь».
Ряд авторов придерживается противоположной точки зрения, считая понятие гомеостаза скорее синонимом сохранительных способностей целостной биосистемы. Клод Бернар, привлекший внимание к стабилизации «внутренней среды», и Уолтер Кэннон, исследовавший гомеостатические функции вегетативной нервной систе мы, черпали силу своих концепций из того, что в наши дни рассматривалось бы как анализ систем»,— писал Уотер-мен [132].
Когда же говорят о поддержании постоянства какой-либо отдельной переменной, например о температурном гомеостазе или гомеостазе сахара крови, то отдают от чет в том, что такой гомеостаз возможен лишь в рамках общего гомеостаза целостной биологической системы.
Как бы ни были хорошо организованы регуляторные механизмы биосистемы, возможности их тем не менее ограничены. Для каждой биосистемы существует область условий внешней среды, в которой эти механизмы и могут обеспечить гомеостаз системы. Температура, влажность, уровень радиации, содержание газов в атмосфере, на личие в окружающей среде пищи и воды — вот некоторые из факторов, определяющих область гомеостаза системы. Для каждого типа биосистем существуют некоторые пре дельные — минимальные и максимальные — значения этих факторов, при выходе за которые система не только не может обеспечить гомеостаз, но и вообще не может под держать стационарное неравновесное состояние.
Вблизи этих границ система может существовать — стационарное равновесное состояние в ней поддерживает ся, но при этом значения существенных переменных ока зываются сильно зависящими от условий внешней среды. И только в средней части допустимой области существо вания биосистемы регулирующие механизмы способны обеспечивать достаточно хороший гомеостаз, т.е. постоян ство значений существенных переменных вне зависимости от внешних условий.
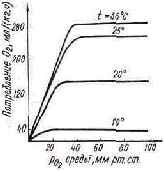
Рис. 3. К определению геометриче ской зависимости: температура тела опоссума ночью ( 1) и днем (2)
Зависимость переменных внутренней среды от внеш них условий для системы, находящейся в стационарном состоянии, при этом при нимает характерный вид кривой с плато посередине и двумя более крутыми участками по краям.
Разумеется, степень вы раженности плато, его ши рина сильно варьируются в разных биосистемах — чем выше организация био системы, тем лучше ее ре гулятивные способности, тем шире плато и тем бо лее полого может идти го-меостатическая кривая.
Типичные гомеостатиче-ские зависимости ряда пе ременных внутренней среды от переменных внешней среды содержат плато, переходящее в обе стороны в более кру тые участки. Положение кривой, вообще говоря, зависит от режима функционирования системы. На рис. 3 приве дена зависимость температуры тела центральноамерикан ского опоссума от температуры окружающей среды ночью и днем. Поскольку это животное ведет ночной образ жиз ни, то в режиме активности (ночью) гомеостатическая кривая располагается выше.
В сравнительной физиологии для описания зависимо стей рассмотренного типа употребляются термины «ре гуляция» и «конформация»: пологие и слабонаклонные участки кривых трактуются как регуляция, прямая пропорциональность—как конформация [112].
На рис. 4 приведен график, иллюстрирующий конформацию и регуляцию темпа потребления кислоро да у животных с независимым дыханием. При снижении давления кислорода в среде рO2 организм увеличивает интенсивность дыхания, сохраняя в некотором диапазоне изменения внешних условий темп потребления О2 неиз менным (регуляция); далее темп потребления кислорода падает (конформация).
Изучение характера гомеостатических кривых, как регуляции, так и конформации, чрезвычайно важно для понимания процессов управления и построения моделей механизмов управления в живых системах.
Рис. 4. Характеристики темпа потребления кислорода.
Большинство живых организмов имеет хорошо выра женные гомеостатические свойства по отношению к ос новным, определяющим фак торам внешней среды. Одна ко имеется и множество форм, не обладающих хорошим го-меостазом вообще. Так, в от ношении теплового режима говорят о гомойотермных и пойкилотермных животных в зависимости от того, насколь ко хорошо выражен у них эффект постоянства темпера туры. Аналогично регуляция водного обмена и осмотиче ского баланса у животных, обитающих в воде, проходит практически все градации —
от пойкилоосмотических животных до гомойосмотиче-ских.
Известно, что гомеостатические свойства организ мов не оказывают существенного влияния на численность вида или ареал их расселения. Исключением, впрочем, являются случаи освоения новых территорий, когда еще не сложилось зрелое экологическое сообщество, и на личия суровых условий обитания (пустыня, тундра). В этих случаях индивидуальные гомеостатические свой ства могут оказаться существенными для данного вида животных. В нормальных же условиях выживание, чис ленность и распространение вида зависят прежде всего от других факторов, например, отношений хищничества, соревнования за жизненные ресурсы [152].
Поэтому можно утверждать, что живые организмы сначала приобрели способность сохранять стационарное неравновесное состояние и лишь затем — свойство го-меостаза. Эта точка зрения принадлежит автору концепции гомеостаза У. Кэннону. «Не предполагается,— пи шет он,— что все гомеостатические механизмы будут найдены у всех форм животных ... Амфибия не способна сохранять содержание воды и постоянство температуры вне зависимости от состояния внешнего мира. Рептилия, высший тип, не так быстро теряет воду через окружающий воздух, как амфибия, но подобно амфибии рептилия хладнокровна и, следовательно, ограничена в своей ак тивности окружающим холодом ... Гомеостаз, наблюдае мый у млекопитающих, является продуктом эволюцион ного процесса» [154]. Можно допустить, следовательно, что первые организмы обладали лишь ограниченными го-меостатическими свойствами [146]. В то же время необхо димое условие существования живых систем — поддер жание стационарного неравновесного состояния или, что то же самое, адекватное снабжение их энергетической системы веществами, выполнялось безусловно.
Единство организма и среды предусматривает непре рывное приспособление организма к изменяющимся условиям как единственно возможный способ существова ния. Каждый акт такого приспособления предполагает возникновение внутри биосистемы некоторых изменений, обусловленных действием внешних сил и противодей ствием организма. В широком смысле под адаптацией ор-_ганизма можно понимать совокупность приспособитель-"ных реакций, протекающих на различных иерархических уровнях в ответ на изменения вещественной, энергети ческой среды и организации воздействующих на организм сигналов. Содержание понятия «адаптация» имеет два аспекта, один из которых связан с процессами приспо собления биосистем к условиям среды, а другой — с со стоянием адаптивности, которое достигается в процессе адаптации [66, 127].
Процесс адаптации биосистемы есть изменение ее биологических (физиологических, биохимических, мор фологических) свойств и поведенческих реакций, направ ленное на сохранение биосистемы как целого и ее гомео стаза на всех системно-структурных уровнях органи зации.
В результате процесса адаптации система приходит в состояние адаптированности, при котором в изменив шихся условиях существования сохраняется обычный (или образуется повышенный) уровень жизнедеятельнос ти и жизнеспособности. Процессы адаптации в биосисте мах протекают с разными скоростями, поэтому иногда различают раннюю адаптацию, которая обеспечивается только адаптационными механизмами биосистемы, и позднюю адаптацию, достижение которой требует вклю чения также и компенсаторных механизмов системы [127].
Состояние адаптации бывает полным, когда в системе сохраняется гомеостаз на всех уровнях организации жиз ни. В этом случае сохраняются и все адаптационно-ком-пенсаторные возможности системы. Неполная адаптация возникает тогда, когда некоторые системы или подсисте мы изменяют свои биологические параметры, но в резуль тате включения компенсаторных механизмов общие, ин тегральные показатели деятельности биосистемы как це лого не нарушаются. При неполной адаптации возможно истощение адаптационно-компенсаторных механизмов, и при изменении условий существования в биосистеме может нарушиться стационарное неравновесное состо яние.
Процесс адаптации возникает в системе всякий раз, когда она попадает в неадекватные условия среды. При этом под адекватными условиями понимаются такие, которые соответствуют генофенотипическим свойствам организма в данный момент [66]. Адаптация к неадекват ным условиям среды требует включения все новых и но вых регуляционных механизмов, с ухудшением условий среды она проходит три стадии — состояние нормы, состояние напряжения механизмов регуляции и, наконец, состояние патологии. Критериями перехода системы от состояния напряжения к патологии являются сохран ность функции и структур и обратимость выявленных отклонений [100, 117].
Одним из важных направлений исследований пробле мы адаптации биосистем является разработка критериев, позволяющих количественно оценивать степень адапта ции, степень напряжения ее механизмов регуляции и пределы адаптации биосистем.
По временным параметрам адаптивные реакции орга низма можно разделить на кратковременные, проявля ющиеся при действии однократных или редко действу ющих раздражений; онтогенетические, проявляющиеся при действии систематических раздражителей в процессе развития и обучения организма; и филогенетические, проявляющиеся в популяции в целом в результате воз никновения у нее на основе изменчивости, наследствен ности и естественного отбора новых биологических ме ханизмов, адекватных новым условиям.
Адаптация может рассматриваться как процесс при способления и как состояние уже достигнутого уровня приспособления к уже изменившимся условиям существо вания [126]. Онтогенетическую адаптацию можно рас сматривать как состояние, промежуточное между нормой и патологией. Истинная адаптация не должна вызывать напряжения защитных и приспособительно-компенсатор-ных механизмов. При этом действие вредных факторов не должно превышать физиологическую меру защиты. По рог вредного действия среды связан с переходными про цессами, лежащими между физиологическими реакциями приспособления и состоянием их «полома» [101]. Состоя ние адаптации, связанное с высоким и постоянным напря жением защитных и приспособительно-компенсаторных механизмов, можно рассматривать как предпатологию, обусловленную неизбежным истощением резервов орга низма.
Особое значение имеет изучение проблемы адаптации организма человека, что связано как с проникновением человека в новую среду обитания (арктические и антарк тические условия, обитание под водой, пребывание в кос мосе), так и с быстрым изменением химического состава среды обитания на Земле.
Процессы адаптации у человека сложнее, чем у живот ных и у других видов биологических объектов. Одним из наиболее существенных отличий является то, что живот ные в процессе адаптации изменяют свои биологические свойства под влиянием условий внешней среды, в то вре мя как человеку свойственно не изменяться при измене нии среды, а создавать условия, которые обеспечивают сохранение гомеостаза на всех системно-структурных уровнях его организма. Точнее говоря, человек не преоб разует среду своего обитания, но включает все большую и большую часть этой среды в состав своеобразной био технической системы, состоящей из организма человека и систем жизнеобеспечения, понимаемых в самом ши роком смысле — от простой одежды и жилища до скафан дров и космических лабораторий.
Биосистемам свойственна динамичность: оператив ная, проявляющаяся в настоящих (в данный момент времени) реакциях на изменения и воздействия окружа ющей среды; онтогенетическая, проявляющаяся в струк турно-функциональных перестройках при развитии биосистемы в онтогенезе при адаптации и обучении; фило генетическая, проявляющаяся в структурно-функцио нальных изменениях биосистемы за счет наследственно сти и эволюции вида.
С момента возникновения (рождения) любая биоло гическая система подвержена изменениям на любом уровне иерархии — от макромолекулярного до поведен ческого. Это свойство обеспечивается различными меха низмами.
Эволюционная динамика (филогенетическая) — му тации, кросинговер, рекомбинации хромосом—направ лена на возникновение новых видов, развитие вида и приспособление его к окружающей среде. Этот вид ди намики связан с перестройкой структур организма и соответствующим изменением функций.
Онтогенетическая динамика выражается в изменении числа клеток, их размеров, размеров органов, развитии систем организма и изменении его форм. На этот вид ди намики оказывает влияние как наследственность, так и воздействие окружающей среды, а изменениям подвер гаются структура и функция. Онтогенетическая динамика проявляется в адаптации всех иерархических уровней организма к изменившимся условиям среды.
Самый быстрый вид динамических изменений парамет ров — это изменения, связанные с конкретной деятельностью организма. Примером могут служить функцио нальные изменения работы сердечно-сосудистой систе мы, системы дыхания при мышечной работе, экстренное изменение двигательной реакции при изменении обста новки.
Для биосистемы характерна качественная неоднород ность, проявляющаяся в том, что в рамках одной и той же функциональной системы совместно и слаженно работают подсистемы с качественно различными адекватными управляющими сигналами (химическими, физическими, информационными).
В качестве вещественных управляющих сигналов вы ступают массы различных неорганических веществ (на пример, ионы, молекулы кислорода, глюкозы), макро молекулы органических гормонов (инсулин, адреналин). Энергетические управляющие сигналы — это теплота, свет, звук, давление, энергия импульсации нейронов. К информационным управляющим сигналам следует отнести, например, словесную обстановку, параметры импульсации нейронных систем (мгновенную частоту, фазу и другие).
Временная неоднородность биосистем проявляется в том, что в одной функциональной системе взаимодей ствуют в достижении одного и того же результата под системы с разными постоянными времени (медленнодей ствующие — биохимические, гормональные; быстродей ствующие — нервная; сверхбыстродействующие — ре чевая, мыслительная, вторая сигнальная).
Практически любая функциональная система орга низма включает все три вида управления — медленно действующее вещественное с постоянной времени, рав ной часам; среднедействующее энергетическое с постоян ной времени, равной минутам, и быстродействующее информационное с постоянной времени, равной секундам.
Временная неоднородность используется организмом при формировании различных способов управления в одной и той же системе с целью достижения оптималь ного результата. Так, на сердечно-сосудистую систему действуют информационные управляющие сигналы сим патических и парасимпатических узлов нервной системы, вещественные — в виде гормонов, энергетические — в виде теплоты, мышечной работы. Все три типа систем управления действуют не изолированно, а взаимосвязан но, что обеспечивает преобразование сигналов различной природы друг в друга.
Взаимодействие биосистем со средой, их постоянное приспособление к среде, эволюция невозможны без ди алектического единства двух противоположных свойств:
структурно-функциональной организованности и струк турно-функциональной вероятностности, стохастичности и изменчивости.
Структурно-функциональная организованность про является на всех иерархических уровнях биосистем и ха рактеризуется высокой устойчивостью биологического вида, его формы и внешнего вида, однообразием составля ющих его элементов, органов и систем. На уровне макро молекул это свойство обеспечивается репликацией макро молекул, на уровне клетки — делением, на уровне особи и популяции — воспроизведением особей путем размножения.
Структурно-функциональная стохастичность био систем функционально проявляется в разнообразии ре акций в ответ на одни и те же воздействия (состояния) сре ды и сигналы биосистем ниже- и вышележащих уровней иерархии. Структурно стохастичность проявляется в способности биосистемы с помощью различных струк турных реализации добиваться однотипного функци онального результата. Наиболее полно это проявля ется при формировании новых условных рефлексов. У биосистем различного уровня иерархии существуют специальные механизмы, обеспечивающие реализацию свойства стохастичности. На уровне макромолекул — это мутации, на уровне клетки — рекомбинации хромо сом, на уровне организма и популяций — рекомбинации генетического фонда.
Динамика стохастичности биосистем проявляется в изменении во времени распределения реакций на один и тот же набор сигналов внешней среды или смежных иерархических уровней.
Важными свойствами биосистем являются также структурная дискретность, без которой невозможно было бы их различие, и функциональная непрерывность, т. е. вариабельность количественных параметров в пре делах одной и той же дискретности.
1.3. ПРИНЦИПЫ ФУНКЦИОНИРОВАНИЯ БИОЛОГИЧЕСКИХ СИСТЕМ
Структурно-функциональные принципы. Эксперимен тальные исследования биологических систем обычно со провождаются теоретическими обобщениями. При этом применение математических методов анализа способству ет более строгому формулированию общих положений.
Ранний период внедрения математических методов в биологию относится ко второй половине прошлого века, когда начала разрабатываться вариационная статисти ка, которую «...Пирсон и его продолжатели специально развивали и расширяли, чтобы сделать ее алгоритмы как можно более пригодными к оценкам, сравнениям и коли чественным характеристикам биологических явлений»
[36].
В начале XX в. П. П. Лазарев, В. Нернст иДж.Леб,
исследуя раздражения нервной ткани, установили коли чественные закономерности и вывели формулы, связы вающие величину возбуждения с силой электрического раздражения. Несколько ранее, в 1901 г., Ж. Вейс пред дожил использовать для этой цели формулу гиперболы. Таким образом, в области нейрофизиологии давно ощу щалось влияние математизации, что сказалось как на проведении самих экспериментов, так и на формулирова нии результатов. Если результаты и не облекались в строгую математическую форму, то формулировались они достаточно логично.
Процесс обобщения результатов и формулирования общих положений, связанных с функционированием био систем, усилился с развитием кибернетики и внедрением в биологические исследования методов моделирования. Общие принципы, выдвинутые кибернетиками при изу чении работы технических систем, оказались вполне применимыми к биологическим.
Рассмотрим группу принципов, связанных со структу рой и функцией биосистем, между которыми существует тесное единство. В каждый момент времени функция формируется на структуре, однако необходимость выпол нять определенную функцию в течение некоторого вре мени неизбежно приводит к формированию новой струк туры.
Одним из общих принципов, связывающих структуру и функцию системы, является принцип наипростейшей конструкции, сформулированный в 1943 г. американским биофизиком Н. Рашевским. Одни и те же функции при одинаковой их интенсивности, вообще говоря, могут вы полняться несколькими различными структурами. Со гласно принципу Рашевского [114] «...та конкретная структура или конструкция, которую мы действительно находим в природе, является простейшей из возможных структур или конструкций, способных выполнять данную функцию или группу функций». В 1954 г. американский ученый Д. Кон усилил формулировку этого принципа и предложил принцип оптимальной конструкции, по которому органическая структура, необходимая для выполнения данной функции, должна быть оптимальной в отношении нужного количества материала и необходи мых затрат энергии. Дальнейшее изучение структур ных особенностей биосистемы привело Н. Рашевского в 1961 г. к принципу адекватной конструкции организма [114]: «Конструкция должна быть адекватной заданной функции при заданных изменяющихся условиях среды».
Сформулированные выше принципы допускают, оче видно, не единственное решение проблемы формы и струк туры. Но это является не недостатком, а преимуще ством. В органическом мире действительно существуют организмы и органы, которые выполняют в основном оди наковые функции, но тем не менее различны по форме. Однако нельзя утверждать, что системы, выполняющие разные функции, всегда различны по своей структуре.
Дополнением к теории адекватной конструкции Ра шевского является принцип эволюционной компенсации силы тяжести [28]. Применительно к рецепторному ап парату вестибулярного анализатора он формулируется следующим образом: в условиях нормальной весомости сила тяжести компенсируется внутренними упругими си лами конструкции рецепторов. В периферической части вестибулярного анализатора в процессе эволюции этот принцип используется для увеличения чувствитель ности рецепторных окончаний специфических нервных клеток.
Обобщением взаимоотношений структуры и функции является принцип структурно-функционального един ства. В любой биологической системе структура и функ ция представляют единое целое, причем функциональный эффект биосистемы достигается за счет ее внутреннего структурирования. Единство структуры и функции био системы формируется в процессе взаимодействия со сре дой и является проявлением адекватности биосистемы среде. В самой биосистеме структура и функция взаимоадекватны.
Принцип структурно-функционального единства при менительно к нервной клетке был сформулирован в 1973 г. [75]: пространственно-временная функциональная не однородность нервной клетки обеспечивается взаимо действием пространственно распределенных структурно неоднородных синаптических образований со структурно однородной мембраной клетки.
Этот принцип справедлив для биосистем любого иерар хического уровня. Чтобы его установить, необходимо детально изучить структурирование биосистемы на под системы и проанализировать формирование функции каж дой подсистемы и биосистемы в целом на основе взаимо действия между подсистемами.
Свойство иерархичности биосистем позволило сфор мулировать принцип этажности [61. В любой сложной системе этажи переработки информации взаимосвязаны и оказывают влияние один на другой. На определенном этапе эволюции биосистем начали возникать этажные структуры. Живые системы усложня лись сначала за счет длины цепей элементов, затем их соединения становились параллельными, далее происхо дила надстройка этажей.
Для клетки этажи — это атомы (если отбросить эле ментарные частицы), простые и сложные молекулы, струк туры из молекул и, наконец, целые клетки. На каждом этаже происходит обмен энергией и вещественными час тицами. В процессе развития каждой регулирующей си стемы организма также формируется сложная структу ра иерархических этажей с вертикальными связями. Од новременно закладываются горизонтальные связи между соответственными этажами близких регулирующих си стем. Для организма прибавляются новые этажи — ткани, органы, системы органов, целостный орга низм.
Затем формируются иерархические уровни систем, объединяющие несколько первичных систем. На каждом уровне сначала образуются параллельные, затем этаж ные структуры. Таким образом, сложные отношения, в которых находятся между собой биологические систе мы организма, носят иерархический характер. Степень независимости одной системы от другой, более крупной, определяется ее жизнеспособностью при отключении от нее энергетических и информационных воздействий со стороны других подобных систем. С понятием сложных отношений связана степень упорядоченности системы или степень непротиворечивости деятельности ее подсистем и элементов, т. е. то, насколько частные функции не ме шают и не противодействуют друг другу.
Повышение степени упорядоченности увеличивает устойчивость системы, но понижает способность ее к эво люции.
Принципы динамического функционирования био систем. Одна из первых попыток формирования принципа работы биосистемы принадлежит П. Мопертюи (XVIII в.). Его принцип наименьшего действия гласит: когда в при роде происходит некоторое изменение, количество дей ствия, необходимое для этого изменения, является наи меньшим возможным. По иронии судьбы принцип Мопертюи, высказанный по отношению к живой природе, к биосистемам, получил строгую трактовку уже безот носительно к биологии, и теперь широко известны егоприложения к теории синтеза технических систем авто матического управления.
Начало анализа сохранительных свойств биосистем связывается с обобщением принципа самосохранения Ле-Шателье: если на систему, находящуюся в устойчи вом равновесии, подействовать извне, изменяя какое-нибудь из условий, определяющих положение равнове сия, то равновесие смещается в том направлении, при ко тором эффект произведенного воздействия уменьшается.
Этот принцип был сформулирован французским хи миком А. Ле-Шателье в 1887 г. для термодинамических систем. После того как немецкий физик К. Браун пока зал, что это положение является следствием второго на чала термодинамики, его иногда стали называть принци пом Ле-Шателье—Брауна.
Аристотель учил, что каждому элементу отвечает свое естественное место во Вселенной, к которому тела стре мятся по природе с тем, чтобы тем достигнуть покоя. После Ньютона, однако, стало ясно, что такого естествен ного места не существует: все тела, если на них действу ют силы, сохраняют свое состояние, а изменение состоя ния, каким бы оно ни было, осуществляется внешними силами. Тела же сопротивляются любому насильствен ному изменению своего состояния.
Б. Спиноза сформулировал свою концепцию самосохранения так: «Никакая вещь не имеет в себе ничего, через' что она могла бы уничтожиться; наоборот, она противо действует всему тому, что может уничтожить ее сущест вование. Следовательно, насколько возможно и насколь ко это от нее зависит, она стремится пребывать в своем существовании» [125].
Принцип Ле-Шателье — Брауна, таким образом, мож но трактовать как частное применение общего принципа самосохранения к термодинамическим системам.
В сходных терминах было сформулировано У. Кен-ноном и понятие самосохранения живых систем: «В от крытой системе, такой как наши организмы, состоящие из нестабильного материала и подверженные непрерыв ному воздействию возмущений, само постоянство служит доказательством существования агентов, действующих или готовых к действию, чтобы поддержать это постоян ство.
Если состояние остается устойчивым, то это происхо дит потому, что любая тенденция к его изменению авто магически вызывает увеличение эффективности фактора или факторов, противодействующих этому изменению» [154].
Важность принципов самосохранения для понимания процессов в живых системах очевидна. Эти принципы в различных сферах исследования используются в различ ных формах. Так, часто принципы сохранения принима ют гомеостатический характер, в той или иной мере близ кий принципу Ле-Шателье [33, 154]. В иных случаях связь концепций с принципом Ле-Шателье отрицает ся [38].
Большое значение для развития представлений об общих принципах работы биосистем имеет сформулиро ванный И. М. Гельфандом и М. Л. Цетлиным принцип наименьшего взаимодействия. Согласно этому принципу, систему можно назвать целесообразно работающей в не которой внешней среде, если система стремится миними зировать взаимодействие со средой [142]. При этом мерой взаимодействия организма со средой может служить от клонение параметров внутренней среды организма от оптимальных значений. Этот принцип был ими выдвинут при изучении механизмов управления движениями и взаимодействием иерархических уровней нервной систе мы. Суть его состоит в том, что если рассматривать слож ную систему управления как совокупность подсистем, составляющих несколько уровней, то каждая такая под система стремится уменьшить свое взаимодействие с внеш ней средой. На каждом уровне внешние воздействия включают идущую с нижних уровней афферентацию, а взаимодействие высших уровней определяет организацию взаимодействия низших уровней. Модель, соответствую щая этому принципу, определяет достижение цели в ме няющейся среде с помощью методов поиска экстремума нестационарной функции многих переменных.
В развитие теоретической биологии большой вклад внес советский биофизик Э. Бауэр, изучивший химиче ские и физические процессы, протекающие в организме на клеточном уровне. Свои выводы он изложил в виде очень оригинальных принципов, облеченных в строгую математическую форму, в книге «Теоретическая биоло гия» (1935 г.). Э. Бауэр подчеркивал, что на уровне за конов химии и физики в организме не наблюдается рав новесия, напротив, организму присуще устойчивое не равновесие. Рассматривая процессы обмена между биосистемой и средой по веществу и энергии, Э. Бауэр [32] формулиру ет некоторые принципы, общие для живых систем:
всем живым существам свойственно прежде всего са мопроизвольное изменение своего состояния, т. е. изме нение состояния, которое не вызвано внешними причина ми, лежащими вне живого организма;
при изменении внешних условий существо не просто противодействует внешней силе (инерции и трению по принципу Даламбера), а в результате противодействия изменяет состояние среды;
работа живых систем при всякой окружающей среде направлена против равновесия, которое должно было бы наступить в данной окружающей среде при данном пер воначальном состоянии системы.
Обобщением этих трех принципов является сформули рованный Э. Бауэром всеобщий закон биологии: все и только живые системы никогда не бывают в равновесии и исполняют за счет своей свободной энергии постоянно работу против равновесия, требуемого законами физики и химии при существующих внешних условиях. Этому закону он придал следующую математическую трак товку:
где F — свободная энергия системы при наступившем равновесии; х— разность факторов работы (разность потенциалов, концентраций); х— изменения, выз ванные этими факторами; l — время, в течение которого произошли эти изменения. Здесь штрих относится к не живой системе, а разность в правой части соответствует работе живой системы против выравнивания.
Практически всему человечеству известны работы И. П. Павлова по условным рефлексам. Здесь мы лишь обсудим соотношение взглядов И. П. Павлова с понятия ми кибернетики. Практически все работы И. П. Павло ва и его школы относятся к исследованию тех или иных форм поведения, к исследованию условий, в которых под влиянием изменившейся окружающей среды разруша ется старый и формируется новый стереотип поведения. Известно также, что в кибернетике принято рассматри вать систему и ее среду и объяснять изменение поведениясистемы изменением среды. С этой точки зрения исследо вание формирования простых, а затем и сложных инстру ментальных рефлексов у животных под действием безус ловных и условных раздражителей есть исследование кибернетических (живых) систем. И результаты, получен ные в этом отношении акад. И. П. Павловым, трудно пе реоценить. Он не использовал математические методы при интерпретации своих экспериментов, но ему удалось сформулировать ряд весьма важных принципов, которые до сих пор не утратили своего значения.
При исследовании взаимодействия организма со сре дой И. П. Павлов обосновал логическую структурную мо дель условных рефлексов. В эксперименте он создал специальные комбинации входных воздействий на ор ганизм, четко регистрировал реакцию организма не толь ко по общему поведению, но и по качественным значени ям физиологических показателей.
Результатом такого структурного подхода явилось формулирование И. П. Павловым в конце 20-х и в нача ле 30-х годов принципа уравновешивания системы со средой, который обобщал три важных положения [104]:
мозг регулирует процессы, протекающие внутри ор ганизма,— принцип нервизма;
мозг устанавливает и регулирует взаимосвязь между организмом и внешней средой на базе безусловных и условных рефлексов — принцип условных и безусловных рефлексов;
кора головного мозга осуществляет функцию образо вания и сохранения динамического равновесия между внутренней средой организма и окружающей организм средой — принцип динамического уравновешивания внешней и внутренней среды.
Если обратиться к истокам кибернетики, нельзя не отдать должное исследованиям И. П. Павлова. И прин цип нервизма, и принцип безусловных и условных реф лексов, и принцип уравновешивания внешней и внутрен ней среды привели к выводу о том, что биологическая система, активно перестраивая свое поведение, стремит ся к уравновешиванию со средой. Это дает ключ к пони манию не только динамического приспособления к изме нившимся условиям среды в онтогенезе, при обучении, но и в эволюции видов, к филогенезу.
Акад. И. П. Павлов является также основателем шко лы, использующей метод логического структурирования биологической модели при исследовании биосистем раз личного иерархического уровня.
Принцип функциональной системы. В 30-е годы идеи, связанные с принципом и теорией функциональной системы, развивал акад. П. К. Анохин. Он придал законченный вид идее рефлекторной дуги И. П. Павлова, характерный для систем с обратной связью. Им был вы двинут и разработан тезис об обратной афферентации— своеобразном замыкании обратной связи организма через окружающую среду — и подчеркнут сложный характер обработки информации в цепи обратной афферентации. Фундаментальным является и положение П. К. Анохина о роли результата как фактора, образующего функцио нальную систему. Как он справедливо отмечает, все опре деления систем, бытующие даже сейчас в кибернетике и общей теории связи, являются неполными вследствие отсутствия связи работы системы с требуемым конечным результатом. Если И. П. Павлов, выдвинув принцип динамического уравновешивания организма со средой, указал на конечную естественную цель, достигаемую ор ганизмом, обитающим в данной среде и приспосаблива ющимся к ней, то теория функциональной системы П. К. Анохина раскрывает, как организм может достиг нуть этой конечной цели, какие механизмы должны для этого действовать. Любопытно отметить, что идеи П. К. Анохина, связанные с анализом биосистем, т. е. наиболее сложных систем, опережают идеи, возникающие в кибернетике на основе анализа и усложнения техни ческих систем, в частности необходимость замыкания об ратной связи обоснована им еще в 1935 г. Обратная аф-ферентация являет собой пример наиболее сложной обработки информации, для которой в технических си стемах нынче используются вычислительные машины;
выдвинутое им положение об акцепторе действия пред восхитило идеи оптимального и критериального управле ния.
Рассмотрим основные положения общей теории функ циональных систем организма. Различные этапы форми рования системы фактически подчинены решению сле дующих вопросов: какой результат должен быть полу чен; когда именно должен быть получен результат;
какими механизмами должен быть получен результат;
как система убеждается в достаточности полученного ре зультата. П. К. Анохин придает результату возможность орга низовать распределение возбуждений в системе в соот ветствующем направлении. Таким образом, все форми рование системы подчинено получению определенного полезного результата; недостаточный результат может целиком реорганизовать систему и сформировать новую, с более совершенным взаимодействием компонентов, даю щим достаточный результат.
П. К. Анохин дает следующее определение понятия системы. [17]: «...системой можно назвать только такой комплекс избирательно вовлеченных компонентов, у ко торых взаимодействие и взаимоотношение приобретают характер взаимосодействия компонентов на получение фиксированного полезного результата».
В свою очередь результат благодаря обратной аф-ферентации имеет возможность реорганизовать систему, создавая такую форму взаимодействия между ее ком понентами, которая является наиболее благоприятной для получения именно запрограммированного результа та. Таким образом, результат рассматривается как не отъемлемый и решающий компонент системы, как ин струмент, создающий упорядоченное взаимодействие меж ду всеми другими ее компонентами.
Функциональные системы организма складываются из динамически мобилизуемых структур в масштабе целого организма. Наличие результата системы как опре деляющего фактора для формирования функциональной системы и наличие специфического строения структурных аппаратов, дающих возможность немедленной мобили зации объединения их в функциональную систему, го ворит о том, что системы организма всегда функциональ ны. Это значит, что функциональный принцип выборочной мобилизации структур является доминирующим. Поэто му такая система и была названа функциональной [15].
С вопросом структурного состава функциональной системы связан и вопрос об иерархии систем. Говоря о составе функциональной системы, следует иметь в виду, что всякая данная функциональная система, взятая для исследования, неизбежно находится где-то между тон чайшими молекулярными системами и наиболее высоким уровнем системной организации.
П. К. Анохин делает следующий вывод о составе ие рархии: все функциональные системы, независимо от уровня своей организации и от количества составляющихих компонентов, имеют принципиально одну и ту же функциональную архитектонику, в которой результат является доминирующим фактором.
Главной чертой каждой функциональной системы яв ляется ее динамичность. Структурные образования, со ставляющие функциональные системы, обладают исклю чительно подвижной мобилизуемостью. Именно это свойство систем и дает им возможность быть пластичными, внезапно менять свою архитектонику в поисках запро граммированного полезного результата.
Применение системы как инструмента в научных ис следованиях затруднено настолько, что многие исследо ватели не изучают внутреннее строение системы, а ограни чиваются исследованием на уровне «черного ящика». Между тем вскрытие внутренних закономерностей дей ствия системы, ее узловых механизмов позволило бы добиться главного в исследовательском процессе: удер жания в руках целого, когда анализируются части этого целого [1б].
Функциональная система всегда гетерогенна. Она состоит из определенного количества узловых механиз мов, каждый из которых занимает надлежащее место и является специфическим для всего процесса формирова ния функциональной системы. Вскрытие этих механиз мов, составляющих внутреннюю архитектонику системы, приблизит исследователей к самой решающей цели системного подхода вообще — обеспечить органическое единство в исследовательском процессе системного уров ня функционирования с индивидуальной характеристи кой каждого дробного элемента или механизма, прини мающего участие в этом функционировании.
Одним из достоинств общей теории функциональных систем является разработка модели системы с четко от работанным внутренним строением или, по выражению П. К. Анохина, внутренней архитектоникой. Такая внутренняя архитектоника, выраженная в физиологи ческих понятиях, является непосредственным инстру ментом для практического применения функциональной системы в исследовательской работе.
Рассмотрим узловые специфические механизмы, пред ставляющие собой внутреннюю архитектонику системы (рис. 5).
Афферентный синтез. Биосистема, даже простой иерархии, сама на основе внутренних процессов прини мает решение о том, какой результат нужен в данный момент ее приспособительной деятельности. Вопрос этот решается именно на стадии афферентного синтеза.
Выдвигаются четыре решающих компонента афферент ного синтеза, которые должны быть подвергнуты одно временной обработке с одновременным взаимодействием на уровне отдельных нейронов: доминирующая на данный момент мотивация; обстановочная афферентация, также соответствующая данному моменту; пусковая афферента ция и, наконец, память
Основным условием афферентного синтеза является
Рис. 5. Общая характеристика функциональной системы (по П. К. Анохину).
одновременная встреча всех четырех составляющих этой стадии функциональной системы. Микроэлектрод ный анализ, микрохимическое исследование и другие формы аналитического исследования нейрона в момент встречи на нем упомянутых выше четырех возбуждений показали, что этот процесс поддерживается и облегчается рядом динамических процессов нервной системы. Это преж де всего выходящая активация, сопутствующая афферент ному синтезу и предшествующая принятию решения [88, 141]. Сюда же относятся процесс корково-подкорковой реверберации [147] и процесс центробежного повышения возбудимости вовлеченных в афферентный синтез ре цепторов.
Афферентный синтез, приводящий организм к реше нию вопроса, какой именно результат должен быть полу чен в данный момент, обеспечивает постановку цели, достижению которой и будет посвящена вся дальнейшая логика системы. Принятие решения является следующим узловым ме ханизмом функциональной системы. Афферентный син тез, подчиняясь доминирующей на данный момент мо тивации, осуществляет подбор тех значений элементов системы, при которых возбуждения избирательно направ ляются к мышцам, совершающим нужное действие. Любое принятие решения после окончания афферентно го синтеза является выбором наиболее подходящих зна чений элементов, которые должны составить рабочую часть системы. Эти значения дают возможность экономно осуществить именно то действие, которое должно привес ти к запрограммированному результату.
Принятие решения — это в высшей степени конденси рованный процесс, в котором одновременно обрабатыва ется на основе доминирующей мотивации вся пришедшая в мозг афферентная информация, производится непрерыв ное сопоставление этих результатов с прошлым ответом и переводятся результаты этой обработки на афферентные пути, соответствующие распределению возбуждений для совершения нужного акта, обеспечивающего получение необходимых результатов.
Формирование акцептора результатов действия. Ак цептор результатов действия является весьма сложным аппаратом. По сути он должен сформировать определен ные нервные механизмы, которые позволяют не только прогнозировать признаки необходимого в данный момент" результата, но и сличать их с параметрами реального ре зультата, информация о которых приходит к акцептору результатов действия благодаря обратной афферента-ции. Именно этот аппарат дает единственную возмож ность организму исправить ошибку поведения или довес ти несовершенные поведенческие акты до совершенных.
П. К. Анохиным с сотрудниками был доказан в ряде экспериментов — ив обычных с условными рефлекса ми, и с помощью тонких электрофизиологических при емов — тот факт, что аппарат акцептора результатов действия организуется непосредственно после принятия решения.
Очевидно, что существенные признаки будущего ре зультата динамически формируются благодаря много сторонним процессам афферентного синтеза с извлечени ем из памяти прошлого жизненного опыта и его результата. Этот комплекс возбуждений является афферентной моделью будущего результата, а именно эта модель, яв ляясь эталоном оценки обратных афферентаций, должна направлять активность человека и животных вплоть до получения запрограммированного результата.
В последнее время были получены сведения о том,что в этот нервный комплекс, обладающий высокой степенью мультиконвергентного взаимодействия, приходит еще одно возбуждение, уже эфферентной природы. Речь идет о коллатералях пирамидного тракта, которые по многим межуточным нейронам отводят «копии» тех эфферент ных посылок, которые выходят на пирамидный тракт. Эти эфферентные возбуждения конвергируют на те же ме жуточные нейроны сенсомоторной области, куда посту пают и все те афферентные возбуждения, которые могут составить параметры реального результата.
Таким образом, момент принятия решения и начала выхода рабочих эфферентных возбуждений из мозга со провождается формированием обширного комплекса воз буждений, состоящего из афферентных признаков буду щего результата и из коллатеральной копии эфферентных возбуждений, вышедших на периферию по пирамидному тракту к рабочим аппаратам. В зависимости от интерва ла между постановкой цели и ее реализацией к этому же комплексу возбуждений через определенное время при ходят возбуждения и от реальных параметров получен ного результата. Сам процесс оценки полученного реаль ного результата осуществляется из сличения прогнози рованных параметров и параметров реально полученного результата. Именно здесь осуществляется оценка по лученного результата. Оценка и результат оценки опре деляют дальнейшее поведение организма. Если результат соответствует прогнозу, то организм переходит к следу ющему этапу поведения. Если же результат не соответ ствует прогнозу, то в аппарате сличения происходит рас согласование, активирующее ориентировочно-исследо вательскую реакцию, которая, поднимая ассоциативные возможности мозга на более высокий уровень, тем самым помогает активному подбору дополнительной информа ции.
Принцип функциональной системы, являющийся до статочно общим, оказался полезным при изучении био систем различного иерархического уровня, а детальное структурирование позволяет проводить эксперименты по выявлению взаимодействия между отдельными блоками системы. Принципы самоорганизации и адекватности. Большое внимание исследователей привлекает такое фундамен тальное свойство биосистем, как способность изменять свою сложность и организацию. Это качество биосистемы изучалось в самых различных аспектах. У. Р. Эшби обратил внимание на необходимость установления соот ветствия между сложностью биосистемы и сложностью среды. В 1958 г. он сформулировал принцип необходимого разнообразия: только разнообразие может уничтожить разнообразие. Этот принцип означает, что биологиче ская система может функционировать в среде, если число состояний системы равно числу состояний среды.
Рис. 6. Схема взаимодейст вия по сложности.
Развитие принципа необходимого разнообразия [25] предусматривает не для всех случаев взаимодействия системы и среды установление равновесия по числу со стояний. Используя меру сложности, введенную У. Р. Эшби, можно записать функции числа состояний для биосистемы и среды:
Нsm = log ns; Hem == log ne
(1.16)
где индекс s относится к систе ме, индекс е — к среде.
Рассогласование между био системой и средой по сложности в этом случае определяется следующим образом:
где kн — коэффициент пропорциональности.
Это рассогласование является управляющим сигна лом, который действует на механизм изменения сложнос ти Wн за счет постоянного притока вещества В и энергии Е (рис. 6).
Таким образом, биосистема, используя устойчивое неравновесие, способна изменить свою структуру и функциональную сложность (число состояний). Слож ность биосистемы изменяется за счет изменения размеров, числа элементов, числа связей между элементами или путем изменения параметров элементов, динамической конфигурации связей между ними. Изменение функци ональной сложности может быть связано с использова нием ранее не задействованных структурных элементов, с изменением порядка и разнообразия реакций биосистемы Измерение числа состояний среды может во многих случаях непосредственно производиться биосистемой. При этом возможны три случая взаимодействия системы со средой в зависимости от величины kн:
1) kн = 1 — каждому состоянию среды соответствует состояние биосистемы — прямое использование принципа необходимого разнообразия Эшби;
2) kн < 1 — каждому состоянию среды может со ответствовать некоторое множество состояний биосисте мы; такой случай возможен, если биосистема обладает возможностью тонкого и структурного анализа состоя ний среды;
3) kн > 1 — определенному набору числа состояний среды соответствует некоторый меньший набор числа со стояний биосистемы; этот случай может соответствовать процессу укрупнения показателей среды по тем или иным критериям, процессу обобщения в образ.
При анализе разнообразия системы и среды нельзя исключить ни один из рассмотренных выше случаев. Более того, найденное тем или иным способом экспери ментальное значение kн может указать на характер вза имодействия биосистемы со средой (анализ, синтез, взаимно-однозначное соответствие). Например, если вза имодействие биосистемы со средой проходит в условиях дефицита вещества и энергии, то в биосистеме может про исходить вынужденное обобщение числа состояний сре ды. Динамика установления необходимого разнообразия в биосистеме может быть различной. Если необходимое разнообразие может быть достигнуто за счет мобилизации структурных и (или) функциональных резервов, то ус тановление может осуществиться достаточно быстро. Если же для необходимого разнообразия биосистема должна заниматься перестройкой собственной структу ры, то этот процесс может быть достаточно длительным. В соответствии с этим основанием для перестройки структуры является частое и длительное воздействие среды на биосистему.
Рассматривая возможности биосистемы по изменению уровня организации, У. Р. Эшби [150] отметил, что само организация как процесс может означать переход от неорганизованной системы к организованной или пере ход от плохо организованной системы к хорошо орга низованной. Он предложил считать критерием само организации биосистем убывание неопределенности, выражаемое условием
В 1960 г. Г. Ферстер ввел в рассмотрение оценку от носительной организации и предложил считать систему самоорганизующейся, если
Усложнение простых условий взаимосвязи биосистемы и среды и учет влияния среды на сложность системы при водят к необходимости рассмотрения текущей неопреде ленности системы как функции распределения вероят ностей состояний. В этом случае условия самоорганиза ции принимают вид [19]:
Рассмотренные выше условия изменения энтропии (не определенности) биосистемы обладают общей следующей особенностью: влияние среды оценивается лишь по изме нению неопределенности или уровня организации био системы.
Более четко принцип самоорганизации был сформули рован в 1962 г. В. М. Глушковым. При этом рассматрива лось взаимодействие системы (в том числе и биосистемы)" со средой и изменение неопределенности системы как функция этого взаимодействия. Пусть Q представляют собой сигналы среды, воспринимаемые биосистемой как обучающая последовательность. Среди всех обу чающих последовательностей есть последовательность Q0, при которой энтропия биосистемы максимальна. Вообще говоря, обучающую последовательность Q0 можно считать тем первоначальным контактом, который биосистема установила с данной средой. Понятно, что если биосис тема способна обучаться, то каждый последующий кон такт со средой может уменьшать неопределенность си стемы. Тогда изменение неопределенности
HQ=HQ-HQ0 (1.21)
является величиной отрицательной.
Часть обучающих последовательностей среды может использоваться или считаться экзаменационными. Пусть биосистема многократно (k раз) может подвергатьсявыражаемое условием
В 1960 г. Г. Ферстер ввел в рассмотрение оценку от носительной организации и предложил считать систему самоорганизующейся, если
Усложнение простых условий взаимосвязи биосистемы и среды и учет влияния среды на сложность системы при водят к необходимости рассмотрения текущей неопреде ленности системы как функции распределения вероят ностей состояний. В этом случае условия самоорганиза ции принимают вид [19]:
Рассмотренные выше условия изменения энтропии (не определенности) биосистемы обладают общей следующей особенностью: влияние среды оценивается лишь по изме нению неопределенности или уровня организации био системы.
Более четко принцип самоорганизации был сформули рован в 1962 г. В. М. Глушковым. При этом рассматрива лось взаимодействие системы (в том числе и биосистемы)" со средой и изменение неопределенности системы как функция этого взаимодействия. Пусть Q представляют собой сигналы среды, воспринимаемые биосистемой как обучающая последовательность. Среди всех обу чающих последовательностей есть последовательность Q0, при которой энтропия биосистемы максимальна. Вообще говоря, обучающую последовательность Q0 можно считать тем первоначальным контактом, который биосистема установила с данной средой. Понятно, что если биосис тема способна обучаться, то каждый последующий кон такт со средой может уменьшать неопределенность си стемы. Тогда изменение неопределенности
HQ=HQ-HQ0 (1.21)
является величиной отрицательной.
По В. М. Глушкову, способность системы к самоорга низации тем выше, чем больше величины S или Z. В отли чие от оценок абсолютной и относительной организации "в данном случае рассматривается изменение неопределен ности не на фоне Нm — максимально возможной неопреде ленности, связанной с внутренними структурными и функциональными возможностями, а на фоне Hm — мак симальной неопределенности, вскрываемой в данной биосистеме определенными воздействиями среды. Такой подход позволяет косвенно через параметры биосистемы судить о свойствах среды и о способности данной биоси стемы обучаться в данной среде.
Таким образом, принцип самоорганизации может по ниматься следующим образом: система обладает способ ностью к самоорганизации в данной среде, если в резуль тате многократного взаимодействия с ней неопределен ность системы по отношению к максимальной неопреде ленности, проявленной системой в данной среде, убывает.
Более прямое сравнение функционирования биосистемы в среде возможно, если рассматривать однотипные пока затели среды и системы; например, сложность Hm и от носительную организацию R. Такой подход позволил сформулировать принцип адекватности биосистемы и сре ды [201. При этом из всей совокупности параметров, по которым биосистема и среда взаимодействуют, были выбраны два наиболее важных: степень сложности и уро вень организации. Для биосистем, действующих в среде постоянной сложности и организации, справедлив статический принцип адекватности: чтобы успешно функ ционировать в среде, сложность и организация биоси стемы должны быть адекватными сложности и органи зации среды. Условно этот принцип можно выразить следующим образом:
где индекс е означает принадлежность к среде, s — к си стеме.
Рассматривая свойства биосистем в связи с функцио нированием их в среде, А. М. Молчанов пишет: «Биоло гические объекты сложны не только внутренне. Они функционируют в сложной, нередко быстро меняющейся среде. Более того, есть серьезные основания думать, что сама сложность их строения носит «компенсаторный» характер. Они именно потому и сложны, что в ответ на любое воздействие среды развивают внешнюю защитную реакцию таким образом, чтобы сохранить максимально неизмененной свою внутреннюю структуру» [92]. Таким образом, «компенсация» воздействий среды есть один из возможных способов установления адекватности между биосистемой и средой.
Установление адекватности между системой и средой по сложности организации является конкретизацией принципа уравновешивания организма со средой И. П. Павлова.
Используем теперь идею И. П. Павлова о динамиче ском уравновешивании применительно к взаимодействию системы и среды. Сформулируем принцип динамической адекватности: при изменении сложности и организации среды биосистема стремится достичь нового уровня аде кватности по сложности и организации со средой с мини мизацией времени, затрат вещества и энергии, т. е.
Hsm(t)Hem(t); Rs(t)Re(t). (1.26)
В динамическом принципе адекватности указывается на оптимизацию таких важнейших параметров жизне деятельности биосистемы, как время, вещество и энергия. При установлении адекватности по сложности и органи зации поведение биосистемы основывается на слож ной форме обратной афферентации и работе акцеп тора действия, а изменение показателей сложности и ор ганизации может подчиняться принципу наименьшего взаимодействия.

Нравится материал? Поддержи автора!
Ещё документы из категории астрономия :
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ