Амитоз, его особенности. Эндомитоз. Политения



Главная » Клетка » Клеточный цикл
Митоз
2n-->S-->4n-->2x2n
ПРОФАЗА.
В профазе происходят следующие события: конденсация хромосом, формирование веретена деления, распад ядрышек, эндоплазматического ретикулума (ЭР), цитоплазматических микротрубочек, снижается и прекращается синтез РНК.
Каждая хромосома двойная (2x2n), они тесно соприкасаются и спирализуются одна относительно другой.
Конденсация хроматина.
После S-фазы сестринские хроматиды остаются связаны мультибелковым комплексом когезинов располагающимся вдоль хроматид в процессе их удвоения. Когезины удерживают хроматиды вместе вплоть до их расхождения в анафазе.
Первый признак Митоза – конденсация хромосом (у человека в 50 раз). Конденсины – белки участвующие в конденсации. Запуск M-Cdk фосфорилирования конденсинов отвечает за их сборку в комплексы на ДНК и конденсации хромосом. При конденсации затрачивается энергия АТФ. Хромосомы конденсируются вокруг продольной центральной оси хромосомы на которой наблюдается наибольшая концентрация конденсинов. В фиксированных препаратах наблюдается сначала спиральная укладка конденсинов вдоль хромосомы (рис.1 )
рис.1 Спиральная укладка хроматид - окраска на специфические белки показывает их спиральное расположение в хромосомах.
Конденсины и когезины структурно родственны и работают по одинаковым механизмам. Установлено, что если после S-фазы соединение хроматид не наступило правильно, то конденсация также не наступает.
Конденсины (когезины) образуют димеры антипараллельно направленные на концах которых находятся ДНК- и АТФ-связывающие домены, а на середине гибкий шарнир (рис.2).
рис.2
Когезины связывают хромосомы еще в S-фазе.
Cohesin is a four-subunit protein complex, in which a heterodimer of SMC proteins, in this case SMC1/SMC3, associates with two other proteins, the Scc1/RAD21/Mcd1 and Scc3 proteins. In vertebrates there are two variants of Scc3, called SA1 and SA2.(Jessberger 2005)
SMC (The structural maintenance of chromosomes proteins) обнаружены в бактериях и археях. В отличии от эукариотических, представляют гомодимеры, кодируемые одним геном.
рис.3 Structure of cohesin and a possible mechanism by which it might hold sister chromatids together. (A) Smc1 (red) and Smc3 (blue) form intramolecular antiparallel coiled coils, which are organized by hinge or junction domains (triangles). Smc1/3 heterodimers are formed through heterotypic interactions between the Smc1 and Smc3 junction domains. The COOH terminus of Scc1 (green) binds to Smc1's ABC-like ATPase head, whereas its NH2 terminus binds to Smc3's head, creating a closed ring. Scc3 (yellow) binds to Scc1's COOH-terminal half and does not make any direct stable contact with the Smc1/3 heterodimer. Scc1's separase cleavage sites are marked by arrows. Cleavage at either site is sufficient to destroy cohesion. By analogy with bacterial SMC proteins, it is expected that ATP binds both the Smc1 and Smc3 heads, alters their conformation, and possibly brings them into close proximity. By altering Scc1's association with Smc heads, ATP binding and/or hydrolysis could have a role in opening and/or closing cohesin's ring. (B) Cohesin could hold sister DNA molecules together by trapping them both within the same ring. Cleavage of Scc1 by separase would open the ring, destroy coentrapment of sister DNAs, and cause dissociation of cohesin from chromatin. (C) Smc-containing complexes other than cohesin could also function via chromatid entrapment. Condensin, for example (black), could organize mitotic chromosomes by trapping supercoils. It and/or other related complexes could hold distant loci together (arrow) and thereby facilitate the function of long-range enhancers and silencers of transcription.
Образование веретена деления
В микротрубочках веретена ~10^8 молекул тубулина. Веретено нормально функционирует при разрушении центриолей лазером. Центром организации микротрубочек служит аморфное вещество центросомы.
Микротрубочки растут от центросом, белки диненины связывают перекрывающиеся микротрубочки, которые продолжают расти и расталкиваются кинезинами, при этом полюса расходятся. В это время микротрубочки с кинетохором не связываются.
Число микротрубочек прикрепленных к кинетохорам различно у разных видов – у некоторых грибов – 1микротрубочка, у человека - 20-40.
Остаточное тельце – фрагменты полюсных микротрубочек+плотный матрикс.
После начала митоза центросомы расходятся и каждая образует радиально симметричный центр организации микротрубочек (астра). Центросома расположена у ядра. Две астры двигаются к противоположным сторонам ядра для формирования двух полюсов веретена деления. Когда ядерная оболочка разрушается (прометафаза) веретено захватывает хромосомы. В клетках эмбрионов Xenopus центросома удваивается даже если ядро было передвинуто, или репликация ДНК подавлена. Центросомный цикл продолжается почти нормально: сначала 2, потом 4, 8 центросом и т.д. На ооцитах Xenopus было показано, что G1/S-Cdk (комплекс cyclin E и Cdk2) инициирует ДНК репликацию в S фазе также стимулирует удвоение центросомы, это предположительно объясняет почему удвоение центросом происходит в начале S-фазы
Рост веретена зависит от моторных белков принадлежащих к двум семействам – kinesin-related proteins движущиеся к ‘+’
концу и денеины, движущиеся к ‘–‘. Три типа микротрубочек наблюдаются в веретене – астральные, кинетохорные, перекрывающиеся-создают правильную структуру веретена. Микротрубочки растут от центросомы вперед ‘+’ концом. Три вида микротрубочек различаются поведением и наборами присоед белков.
Веретено начинает собираться в профазе. M-Cdk запускают фосфорилирование двух типов белков контролирующих динамику микротрубочек. Типы: моторные белки и microtubule-associated proteins (MAPs). Также имеются белки катастрофины.
В интерфазе микротрубочки отходят от одной центросомы и находятся в динамическом равновесии. Переключение ведущее к росту называется спасение, переключение к уменьшению микротрубочек – катастрофа. В профазе длинные интерфазные микротрубочки быстро преобразуются в множество коротких окружающих каждую центросому, которые начинают формировать веретено деления.
РАСПАД ЭР
ЭР распадается на мелкие вакуоли, лежащие по переферии клетки и Аппарата Гольджи (АГ), который теряет околоядерную локализацию, разделяется на отдельные диктиосомы разбросанные в цитоплазме.
ПРОМЕТАФАЗА
Распад ядерной оболочки, беспорядочное движение хромосом в области бывшего ядра, хромосомы через кинетохор соединяются с веретеном и начинают движение.
Распад ядерной оболочки
Кинетохор
Sc: кинетохор связан с цетромерным локусом CEN: CDEI,II,III. CDEI,III – консервативные районы сходны с Dm. CDEII – обогащен АТ, участок разной длины. CDE ответственен за связь с мт, взаимодействует с рядом белков.
кинетохор – мультибелковый комплекс, состоит из трех слоев:
наружный – плотный (СENP-E, СENP-F – участвуют в связывании мт), от него отходит множество фибрилл – фиброзная корона кинетохора (СENP-E, динеины)
средний – рыхлый, 3F3/2 – белок, регистрирует натяжение пучков мт
внутренний – плотный, участок ГХ обогащенный а-сателлитной ДНК (СENP-B- связывается с а-ДНК, MCAK-кинезинподобный белок-когезин, INCENP-когезин, СENP-А-аналог H3, СENP-G-связывается с белками ядерного матрикса, СENP-С-ф-ция не выяснена)
Функция кинетохора: связывание хроматид, закрепление мт веретена.
min число мт у Sc 1 на хромосому, у высших растений 20-40 мт на хромосому
белки кинетохора присутствуют во всех стадиях кц, образование и деление кх происх в S-периоде
Х-мы беспорядочно движутся – метакинез – то приближаются к полюсам, то удаляются к центру веретена, пока не займкт среднее положение – конгрессия х-м. мт случайно захватываются кинетохором и х-мы скользят по мт к полюсу 25мкм/мин, с помощью аналога динеина. Во время движения мт не разбираются. Хроматиды связаны и тянутся с двух сторон. Если лазером перерезать мт с одной стороны, то х-мы утянуться к противоположному полюсу
Перемещение хромосом к экватору
если митотич кл обработать D2O или таксолом – подавляют разборку мт?мт удлиняются и не тянут хромосомы?блок митоз
колхицин, низкая t, высокое гидростатич давление – разрушение нитей веретена?блок митоз
сила действующая на кинетохорную нить тем слабее чем ближе к полюсу нах кинетохор
МЕТАФАЗА
Завершается формирование веретена деления, хромосомы перестают двигаться и выстраиваются по экватору веретена (экваториальная пластинка)
метафаза - синтез белка – 20-30% от интерфазы. Клетки наиболее чувствительны к холоду, колхицину и др. агентам, которые разрушают веретено деления и приводят к прекращению митоза (К-митоз), при малых дозах митоз восстанавливается через несколько часов (иначе гибель либо полиплоидия).
Метафаза – хромосомы образуют пластинку, микротрубочки достигают max концентрации и перекрываются.
АНАФАЗА
Анафаза – хромосомы внезапно одновременно отделяются друг от друга и начинают движение к полюсам. Центромеры разъединяются – деградация центромерных когезинов. Наиболее короткая стадия, разделение хроматид и расхождение хромосом к полюсам (v=0,2-5 мкм/мин). Иногда также расходятся полюса друг от друга.
Расхождение хромосом за счет кинетохорных пучков микротрубочек – анафаза А, расхождение хромосом вместе с полюсами за счет удлинения межполюсных микротрубочек – анафаза В.
Разделение хроматид и движение к полюсам.
Веретено и перетяжка связаны так, что пока хромосомы не разойдутся перетяжка цитоплазмы не наступает.
События анафазы: движение кинетохорных нитей к полюсам, движение полюсных нитей расталкивающих полюсы-движутся друг относительно друга; малые дозы хлоралгидрата предотвращают удлинение и движение полюсных нитей, но не влияют на кинетохорную нить.
ТЕЛОФАЗА.
Телофаза длится с момента прекращения движения хромосом. Происходит реконструкция ядер - образование ядерной оболочки, деспирализация хромосом, активация хромосом - увеличение уровня транскрипции, формирование ядрышек, разрушение веретена деления, разделение клеток, образование остаточного тельца Флеминга, образование перетяжки.
В местах контактов хромосом с мембранными пузырьками начинает образовываться ядерная оболочка. Сначала она образуется на латеральных поверхностях хромосом, затем в центромерных и теломерных участках. После смыкания ядерной оболочки происходит образование ядрышек.
Разборка микротрубочек идет от полюсов к экватору бывшей клетки, в средней части веретена микротрубочки сохраняются дольше всего – остаточное тельце.
Цитокинез.
Борозда деления образуется в плоскости метафазной пластинки под прямым углом к длинной оси митотического веретена. Перетяжка содержит актиновые филаменты и миозин II, расположенные по экватору делящейся клетки под плазматической мембраной (ПМ) стягивая ее изнутри.
Одной из причин почему цитокинез не происходит раньше окончания митоза является активность M-Cdk инактивируемой в конце митоза.
ИНТЕРФАЗА.
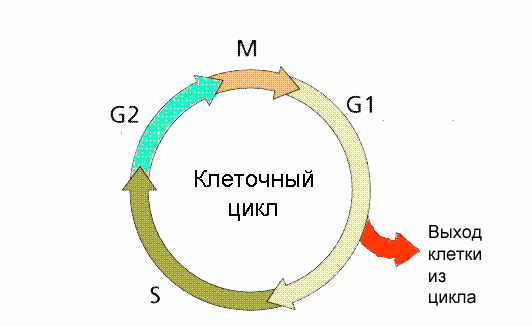
КЛЕТОЧНЫЙ ЦИКЛ
Функция воспроизведения и передачи генетической информации обеспечивается в ходе клеточного цикла.
Клеточный цикл - совокупность явлений между двумя последовательными делениями клетки или между ее образованием и гибелью Клеточный цикл включает собственно митотическое деление и интерфазу - промежуток между делениями.
Интерфаза
Интерфаза значительно более длительна, чем митоз (обычно занимает не менее 90% всего времени клеточного цикла) и подразделяется на три периода: пресинтетическип или постмитотический (G1), синтетический (S) и постсинтетический или премитотический (G2).
1. Пресинтетический или постмитотический (G1) период (от англ. gap - промежуток) наступает сразу же после митотического деления клетки и характеризуется активным ростом клетки и синтезом белка и РНК, благодаря чему клетка достигает нормальных размеров и восстанавливает необходимый набор органелл. G1 -период длится от нескольких часов до нескольких дней. В течение этого периода синтезируются особые "запускающие" белки (trigger proteins), или активаторы S-периода. Они обеспечивают достижение клеткой определенного порога (точки R - рестрикции или ограничения), после которого она вступает в S-период.
Контроль, осуществляемый на уровне точки R (при переходе из G1 в S), ограничивает возможность нерегулируемого размножения клеток. Проходя эту точку, клетка переключается на последующую регуляцию внутренними факторами клеточного цикла, которая обеспечивает закономерное завершение ее деления.
Если клетка не достигает точки R, она выходит из цикла и вступает в период репродуктивного покоя (G0) для того, чтобы (в зависимости от причин остановки): (1) дифференцироваться и выполнять свои специфической функции, (2) выжить в условиях недостаточности питательных веществ или факторов роста, (3) осуществить репарацию поврежденной ДНК. Клетки одних тканей при соответствующей стимуляции вновь способны возвращаться из периода (G0) в клеточный цикл, других - утрачивают эту способность по мере дифференцировки
2. Синтетический (S-) период характеризуется удвоением содержания (репликацией) ДНК и синтезом белков, в частности, гистонов, которые поступают в ядро из цитоплазмы и обеспечивают нуклеосомную упаковку вновь синтезированной ДНК. В результате происходит удвоение числа хромосом. Одновременно удваивается число центриолей. S-период длится у большинства клеток 8-12 часов.
3. Постсинтетический или премитотический (G2-)период следует за S-периодом и продолжается вплоть до митоза (часто обозначаемого буквой М). В течение этого периода клетка осуществляет непосредственную подготовку к делению. Происходит созревание центриолей, запасается энергия, синтезируются РНК и белки (в частности, тубу-лин), необходимые для процесса деления. Длительность G2-периода составляет 2-4 часа. Возможность выхода клетки из G2-периода в G0-neриод с последующим возвращением в G2-период в настоящее время большинством авторов отрицается.
Контроль вступления клетки в митоз осуществляется двумя специальными факторами с противоположно направленными эффектами. Митоз тормозится до момента завершения репликации ДНК М-задерживающим фактором и индуцируется М-стимулирующим фактором. Действие последнего проявляется лишь в присутствии других белков - циклинов (синтезируются на протяжении всего цикла и распадаются в середине митоза).
Регуляция клеточного цикла
По уровню обновления клеток все ткани организма подразделяются на три группы.
(1) стабильные клеточные популяции - состоят из клеток с полной потерей способности к делению (нейроны, кардиомиоциты). Число клеток в такой популяции стабилизируется в начале их дифференцировки; по мере старения организма оно снижается вследствие невосполняемой естественной убыли клеток.
(2) растущие клеточные популяции способны не только к обновлению, но также и к росту, увеличению массы ткани за счет нарастания числа клеток и их полиплоидизации. Их долгоживущие клетки выполняют специализированные функции, но сохраняют способность при стимуляции вновь вступать в цикл с тем, чтобы восстановить свою нормальную численность. Описанные популяции клеток образуют почки, печень, поджелудочную и щитовидную железы
(3) обновляющиеся клеточные популяции характеризуются постоянным обновлением клеток; убыль
дифференцированных, выполняющих специализированные функции и неспособных к делению клеток вследствие их гибели уравновешена образованием новых в результате Деления малодифференцированных камбиальных клеток и их последующей дифференцировки. К таким популяциям относят эпителий кишки, эпидермис, а также клетки костного мозга и крови Регуляция клеточного цикла в различных тканях организма осуществляется сбалансированной сложной системой механизмов, стимулирующих или ингибирующих клеточное деление. Система регуляции клеточного цикла получает два вида информации.
(1) о действии на клетку различных внешних факторов, способствующих активации или торможению ее деления Она обрабатывает и интегрирует ее в виде сигналов, определяющих, будет ли клетка вступать в митотический цикл или дифференцироваться и пребывать в периоде репродуктивного покоя (G0).
(2) об интактности генома. При повреждении генома клетки прохождение ею цикла останавливается и включается система репарации ДНК. Тем самым снижается вероятность нежелательной репликации поврежденной ДНК. Многочисленные сигналы, регулирующие деятельность клетки, замыкаются на ген р53, который блокирует прохождение клеточного цикла до устранения возникшего повреждения. Если это повреждение слишком серьезно, р53 (в совокупности с другими регуляторами) запускает программу апоптоза - запрограммированной гибели клетки

Нравится материал? Поддержи автора!
Ещё документы из категории биология:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ