Будова, функції та методи дослідження мітохондрій

 ЗМІСТ
ЗМІСТ
ВСТУП
РОЗДІЛ 1. МІТОХОНДРІЇ ЯК ОРГАНОЇДИ КЛІТИНИ, ЇХ БУДОВА ТА ФУНКЦІЇ
1.1 Загальна будова мітохондрій
1.2 Генетична система мітохондрій
1.3 Функції мітохондрій
1.4 Походження мітохондрій
РОЗДІЛ 2. МЕТОДИ ДОСЛІДЖЕННЯ МІТОХОНДРІЙ
РОЗДІЛ 3. ДОСЛІДЖЕННЯ МІТОХОНДРІЙ НА СУЧАСНОМУ ЕТАПІ
ВИСНОВКИ
СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
ВСТУП
Мітохондрії (від греч. mitos - нитка і chondrion - зернятко, крупинка), хондріосоми, які постійно присутні в клітинах тварин і рослин, органоїд, який забезпечує клітинне дихання, у результаті якого енергія вивільнюється або акумулюється в легко використовуваній формі. Вони відрізняються рядом характерних морфологічних, біохімічних та функціональних властивостей, до яких можна віднести розмір, форму, особливість зафарбовуватись, специфічну структурну організацію, ліпопротеїдний склад та вміст великої кількості ферментів і коферментів, які разом приймають участь в процесах трансформації енергії в клітині. В фізіологічному відношенні мітохондрії являють собою біохімічну „машину”, призначення якої видобувати енергію з харчових речовин та акумулювати її за допомогою фосфорилювання в фосфатних зв’язках аденозинтрифосфата. Таким чином, мітохондрії – органоїди, які продукують енергію життєво необхідну для клітини.
Значних успіхів в дослідженні мітохондрій було досягнуто з використанням таких методів, як електронна мікроскопія та біохімічний аналіз цих органоїдів після їх виділення. Особливо багато було зроблено в 50-60-х рр. минулого століття. Тому виникає непідробний інтерес до з’ясування ролі мітохондрій, особливостей їх функціонування та успадковування, їхньої специфічної генетики.
Метою нашої курсової роботи ми обрали визначення досягнень сучасної біології у тій області, яка стосується вивчення мітохондрій. Відповідно до мети потрібно вирішити такі задачі:
на основі літературних джерелах встановити будову мітохондрій та їх функції
з’ясувати особливості мітохондріального геному, його функціонування, процеси успадковування;
визначити методи, які використовуються при дослідженні мітохондрій як субклітинних структур;
провести аналіз сучасних досліджень з даного питання на основі матеріалів журналу „Цитологія и генетика”.
Огляд сучасних матеріалів з цієї теми дасть можливість оцінити стан та глибину досліджень, які проводяться в цій царині нашими науковцями.
РОЗДІЛ 1. МІТОХОНДРІЇ ЯК ОРГАНОЇДИ КЛІТИНИ, ЇХ БУДОВА ТА ФУНКЦІЇ
1.1 Загальна будова мітохондрій
Вивчення біохіміками і цитологами мітохондрій лише в 40-50-х роках нашого сторіччя, коли методами диференціального центрифугування вдалося одержати чисті фракції мітохондрій і показати, що в них локалізуються ферменти дихального ланцюга, циклу Кребса й окисного фосфорилювання; у 1952-53 р. були проведені дослідження ультраструктури мітохондрій.
Мітохондрії відсутні у прокаріот (бактерій, синьозелених водоростей) і зустрічаються практично у всіх еукаріотичних клітинах, за винятком деяких паразитичних протестів (Entamoeba hystolytica, Trychomonas) і вільноживучої Реlomyxa polustris, що живе в умовах дефіциту кисню. Кількість мітохондрій у клітині значно варіює: клітки деяких водоростей і містять по одній мітохондрії, сперматозоїди різних видів тварин - від 20 до 72, у соматичних клітинах ссавців, як правило, 500-1000 мітохондрій, а в гігантської амеби Chaos chaos їхнє число досягає 500 000.
Однак об'ємні реконструкції мітохондрій по серіях зрізів, отримані для багатьох об'єктів за допомогою електронно-мікроскопічних досліджень, показали, що кількість мітохондрій у клітинах, очевидно, значно менша, ніж передбачалося раніш.
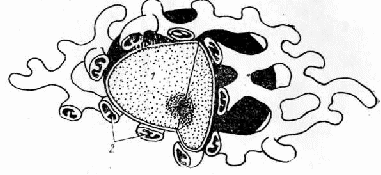
Так, у дріжджів, грибів, у гаметах хламідомонад є одна гігантська, сильно розгалужена мітохондрія. Природно, що на звичайних зрізах при цьому виявляються багаточисленні дрібні мітохондрії (рис. 1.1). У фібробластах і нервових клітках мозку, які культивуються in vitro, при реконструкції виявляються довгі нитковидні мітохондрії, а в лімфоцитах - дві-три великі мітохондрії, розташовані навколо ядра. У поперечносмугастих м'язових волокнах невелика кількість сильно розгалужених гігантських мітохондрій утворює так звану мітохондріальну мережу.
Рис. 1.1 - Гігантська мітохондрія гриба: 1 – ядро, 2 – ділянки мітохондрій в поперечному розрізі
Пластичність організації мітохондрій виявляється в їх змінах в онтогенезі. Наприклад, спочатку в зооспорі одного з ооміцетів всього одна велика мітохондрія. В ході розвитку вона стає більш тонкою, у ній з'являються наскрізні отвори і в кінці утворюються чотири мітохондрії: одна велика і три дрібні. В окремих представників голкошкірих велика частина дрібних мітохондрій в ооцитах по мірі росту цих клітин зливаються, і виникає гігантська кільцеподібна мітохондрія, яка оточує звичайні дрібні мітохондрії і деякі інші органоїди (зокрема, апарат Гольджі). Після попадання ооцитів у морську воду відбувається відгалуження, а потім і фрагментація гігантської кільцеподібної мітохондрії на численні дрібні. Одночасно з цим спостерігається активізація дихання ооциту. Подібні зміни відмічаються в ході клітинного циклу у Euglena gracilis. Дрібні мітохондрії об’єднуються в гігантську, котра потім фрагментується. І в цьому випадку активізація дихання збігається з утворенням численних дрібних мітохондрій з однієї гігантської.
Розміри і форма мітохондрій, які виявляються на звичайних ультрамікроскопічних зрізах, сильно варіюють. За формою вони бувають нитковидними, паличковидними, округлими і гантелеподібними навіть у межах однієї клітки. Крім того, фазово-контрастна мікроскопія живих кліток показала, що мітохондрії - дуже динамічні структури: вони можуть рости в довжину, стискуватися, розгалужуватись, ділитися - і все це швидше, ніж за одну хвилину.
Мітохондрії розташовуються в клітині, як правило, або в тих ділянках, де витрачається енергія, або біля скупчень субстрату (наприклад, ліпідних крапель). Сувора орієнтація мітохондрій виявляється вздовж джгутика у сперматозоїдів; в епітеліальних клітинах ниркових канальців мітохондрії локалізуються в складках базальної мембрани, утворюючи разом з нею апарат активного транспорту іонів, який характерний видільному і осморегулюючому епітелію. Скупчення мітохондрій виявляється в області синапсів і фоторецепторних клітин у основи зовнішнього членика, де вони складають особливу структуру - міоїд. Природно, що таке розташування мітохондрій зменшує втрати АТФ під час її дифузії.
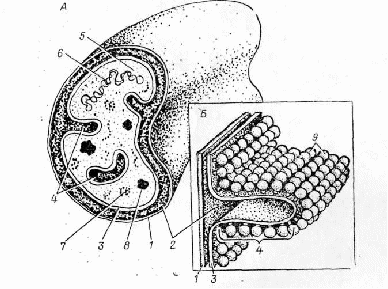
Як показано на великому порівняльно-цитологічному матеріалі, загальний план будови мітохондрій однаковий у всіх еукаріот. Мітохондрії всіх організмів від дріжджів до вищих тварин оточені двома мембранами, між якими розташовується міжмембранний простір. Внутрішня мембрана утворює вирости в мітохондріальний матрикс - кристи; у матриксі містяться ДНК, рибосоми і різні включення. Рибосоми іноді прикріплюються до внутрішньої мембрани або утворюють полісомні ланцюги.
Форма крист може бути пластинчастою або трубчастою, вони розташовуються або паралельно довгій осі мітохондрії (аксони нервових клітин, поперечно-смугасті м’язи), або перпендикулярно їй (клітини печінки, нирок). Кристи мітохондрій досить лабільні утворення і легко переходять з однієї форми в іншу, а іноді можуть і редукуватися. Наприклад, при анаеробному розвитку дріжджів кристи майже зникають; якщо знову створити аеробні умови, то кристи відновлюються.
Структура мітохондрій залежить від функціональної активності тканини й організму. При цьому може змінюватися не тільки форма і кількість мітохондріальних крист, але і кількість самих мітохондрій. У функціонально аналогічних структурах у филогенетично віддалених тварин мітохондрії схожі між собою більше, ніж у різних органах одного і того ж виду. Наприклад, у структурі мітохондрій літальних м'язів голуба і бабки більше подібності, чим між мітохондріями літальних і скелетних м'язів бабки (рис. 1.2).
Рис. 1.2 - Загальна схема організації мітохондрій (А) та ділянка кристи з грибоподібними тілами (В): 1 – зовнішня мембрана, 2 – міжмембранний простір, 3 – внутрішня мембрана, 4 – кристи, 5 – мітохондріальний матрикс, 6 – ДНК, 7 – рибосоми, 8 – конкреції фосфату кальцію, 9 – грибовидні тіла
Кількість і ступінь розвитку крист визначаються функціональною активністю тканини. Так, у мітохондріях спороцист Fasciola hepatica, паразита печінки молюсків, кристи, одиничні. У церкарії, яка веде активний вільний спосіб життя, число крист дуже велике. Мітохондрії більшості рослинних клітин звичайно мають мало крист, але в секреторних клітинах рослин їхній так само багато, як і у тваринних клітинах.
Мітохондрії - лабільна структура, тому вони легко піддаються адаптивним перебудовам. Так, при гіподинамії у пацюків кількість як крист у мітохондріях, так і самих мітохондрій у поперечно-смугастих м'язах різко зменшується. Якщо ж цих тварин змусити активно рухатися (наприклад, плавати), мітохондрії настільки ж швидко приймають колишній вид, і відновлюється їхня вихідна кількість.
Зовнішня і внутрішня мембрани мітохондрій значно розрізняються між собою. При підвищенні або зниженні осматичного тиску внутрішня мембрана відповідно зморщується або розправляється, легко переходячи з одного стану в інше; зовнішня мембрана здатна лише до необоротнього розтягнення, яке веде до розриву. Видаляючи таким чином зовнішню мембрану, одержують препарати ізольованої внутрішньої мембрани мітохондрій. Відрізняються мембрани і проникливістю: зовнішня мембрана характеризується неспецифічною проникливістю, а проникність внутрішньої, навпроти, високоспецифічна. Мембрани мітохондрій неоднакові і по стійкості до різних ферментів і детергентам.
Розбіжності у властивостях мітохондріальних мембран обумовлені значними розбіжностями в їхній структурі. Так, співвідношення ліпідів і білків у зовнішній мембрані складає 0,88 (тобто до її складу входить менше 20% білка), а у внутрішній - 0,30 (75% білка). Ліпіди внутрішньої і зовнішньої мембран розрізняються зо вмістом насичених жирних кислот - у ліпідах внутрішньої мембрани їх більше. Неоднаковий і склад ліпідів у мембранах. Так, у внутрішній мембрані дуже низький рівень вмісту холестерину і високий - кардіоліпіну, „подвійного” фосфогліцериду, який складається з двох залишків фосфатидної кислоти, зв'язаних один з одним гліцерином. Цей ліпід практично не зустрічається в інших мембранах еукаріотичних клітин і, очевидно, обумовлює малу проникливість внутрішньої мітохондріальної мембрани для іонів.
Неспецифічна проникникливість зовнішньої мембрани визначається наявністю в ній білків поринів, які формують у ліпідному бішарі численні канали, через які можуть проходити молекули масою до 10 кДа.
Зовнішня мембрана мітохондрій бідна ферментами; небагато їх і в міжмембранному просторі. Зате внутрішня мембрана і мітохондріальний матрикс буквально насичені ними. Так, у матриксі зосереджені ферменти циклу Кребса й окислювання жирних кислот. У внутрішній мембрані локалізований ланцюг переносу електронів, дихальний ланцюг, ферменти фосфорилювання і численні транспортні системи, що забезпечують її виборчу проникність. Мітохондрії являють собою структури, які здатні обновлюватися та мають досить короткий життєвий цикл (у клітках печінки пацюка, наприклад, період напівжиття мітохондрій складає близько 10 днів). Мітохондрії утворюються в результаті росту й поділу поппередньо існуючих мітохондрій. Цей процес іноді протікає дуже швидко і з великою інтенсивністю (зокрема при адаптивних перебудовах). Діляться мітохондрії за допомогою перетяжки.
1.2 Генетична система мітохондрій
Мітохондрія має власну генетичну систему - мітохондріальну ДНК і власний білоксинтезуючий апарат. Діленню (репродукції) мітохондрій передує реплікація мітохондріальної ДНК. Синтез ДНК мітохондрій при діленні клітин відбувається незалежно від реплікації ядерної ДНК.
У більшості об'єктів ДНК мітохондрій являє собою дволанцюгову ковалентно замкнуту структуру. В одній мітохондрії може бути від однієї до 20 000 кільцевих молекул ДНК, молекулярна маса яких варіює від (9-10) • 106 у хребетних до (3-4) • 107 у вищих рослині і грибів. Розміри кільцевих молекул ДНК у більшості мітохондріальних геномів варіюють від 16-20 (у хребетних) до 570 кілобаз у вищих рослин (кукурудза), а лінійна довжина молекули складає 5 мкм у дріжджів і 30 мкм у вищих рослин.
У мітохондріях тваринних клітин знаходиться кілька молекул ДНК, практично не помітних на звичайних електронно-мікроскопічних препаратах. У клітинах рослин їх, як правило, більше, так що при ультраструктурних дослідженнях мітохондріальна ДНК може бути виявлена у виді тонких фібрил. І, нарешті, у мітохондріях деяких кінетопластид (наприклад, трипаносомових) безліч кільцевих молекул ДНК поєднуються у велику електронно-щільну структуру - кінетопласт. У єдиній гігантській мітохондрії кінетопластид виявляється цікава модифікація мітохондріального геному: генетичний апарат тут представлений великою кількістю кільцевих молекул двох типів - так званих максі-кілець (вони гомологічні кільцевим ДНК мітохондрій інших об'єктів) і мінікілець - вони майже не містять генетичної інформації і їхнє функціональне значення стало прояснятися лише останнім часом. У рідких випадках, наприклад у водорості хламідомонади і інфузорій парамецій, мітохондріальний геном може складатися з лінійних молекул ДНК.
Про надмолекулярне упакування мітохондріальної ДНК відомо небагато. Очевидно, фібрили мітохондріальної ДНК за способом упакування нагадують хромосому прокаріот. У деяких об'єктів виявлені основні білки невеликої молекулярної маси, начебто зв'язані з мітохондріальною ДНК. Мітохондріальна ДНК відрізняється від ядерної за складом основ, плавучій щільності і ряду інших фізико-хімічних властивостей.
Значних успіхів досягла генетика мітохондрій. Численні роботи проводяться на мітохондріальних мутантах дріжджів (мутації дихального ланцюга, які не ведуть до загибелі кліток, тому що вони переходять на бродильний тип обміну). При дослідженнях клітин, які культивувалися in vitro вдалося одержати численні мітохондріальні мутанти в птахів, ссавців (у тому числі і людини); були отримані навіть клітини з мітохондріями без власної ДНК.
Дослідження в області мітохондріальної генетики з використанням мутантних клітин дозволили встановити, що ДНК мітохондрій властиві всі генетичні функції: рекомбінація, репарація і т.д. Було показано, що мітохондрії характеризуються так називаною полярністю, тобто перевагою в мітохондрій дочірніх клітин рекомбінантів одного типу: успадковування мітохондріального геному у тваринних і рослинних клітин в основному іде по материнській лінії. Правда, ці дані були отримані на обмеженому числі об'єктів. Останнім часом були виявлені факти спадкування обох типів батьківських мітохондріальних геномів у тваринних клітин і спадкування тільки по батьківській лінії в рослинних клітинах (секвойя). Для позначення цього явища - наявності в індивіда більше одного класу мітохондріального генома (або генома хлоропластів) запропонований спеціальний термін - гетероплазмія.
Дослідження в області мітохондріальної генетики дозволили картувати мітохондріальні геноми в різних об'єктів. Зокрема, є повна характеристика геномів мітохондрій клітин людини і дріжджів; про інші організми існують уривчасті відомості.
Обсяг інформації, закодованої в геномі мітохондрій, відносно невеликий; мітохондріальний геном, як правило, містить матриці для синтезу власних тРНК і високомолекулярних рРНК і деяких субодиниць ферментів дихального ланцюга й АТФ-синтазного комплексу.
Автономність білоксинтезуючої системи мітохондрій значно менша, ніж здається на перший погляд. Білкові фактори трансляції мітохондріальних РНК кодуються не ДНК мітохондрій, а ядерною ДНК, тобто процес трансляції в мітохондріях не може здійснюватися незалежно від ядерного геному. Крім того, у ДНК ядра є інформація, необхідна для синтезу факторів реплікації мітохондріальної ДНК, РНК-полімераз, що здійснюють транскрипцію ДНК мітохондрій, білків, які входять до складу 70 S рибосом (за винятком одного білка малої субодиниці) і більшої частини компонентів електроннотранспортного ланцюга й АТФ-синтазного комплексу.
В даний час велика увага приділяється вивченню механізмів транслокації білків через мітохондріальні мембрани. Встановлено, що численні білки дихального ланцюга, АТФ-синтазного комплексу і матриксу проходять у мітохондрії, очевидно, у ділянках тісного контакту зовнішньої і внутрішньої мембран - так називаних контактних сайтах. Тут знаходиться спеціальний білок-рецептор молекулярною масою 45 кДа, і утворюються специфічні канали. Процес цей вимагає енергії АТФ.
Більшість мітохондріальних мембранних ліпідів синтезується на ЕПС і вбудовується в мембрану мітохондрій. Ліпіди надходять до зовнішньої мембрани мітохондрій і попадають у внутрішню мембрану через контактні сайти. У мітохондріальних мембранах відбувається перетворення екзогенного фосфатидилсерину у фосфатидилетаноламін і утворення з «імпортованих» ліпідів кардіоліпіну - специфічного ліпіду внутрішньої мембрани мітохондрій.
Дослідження геному і білоксинтезуючого апарату мітохондрій дозволили прийти до висновку, що їм властиві деякі специфічні риси. Так, у мітохондріях має істотні особливості їх генетичний код. Універсальний термінінуючий кодон UGA у мітохондріях ссавців, дрозофіл і дріжджів кодує триптофан, а ізолейциновий кодон AUA - метіонін. Більш того, мітохондріальний генетичний код відрізняється в різних організмів. Так, лейциновий кодон GUA у мітохондріях ссавців і дрозофіл також кодує лейцин, а в мітохондріях дріжджів - треонін. Кодони AGA і AGG, які несуть в універсальному коді інформацію для синтезу аргініну, функціонують як термінуючі кодони в мітохондріях ссавців, кодують серин в мітохондріях дрозофіл і аргінін у мітохондріях дріжджів. У вищих рослин мітохондріальний генетичний код збігається з ядерним. Для зчитування всіх кодонів у білоксинтезуючому апараті мітохондрій досить всього 22-24 тРНК на відміну від цитозолю (де є принаймні 31 тРНК) і хлоропластів (30 тРНК); трансляція в мітохондріях відбувається з меншою точністю; мітохондріальні рибосоми також мають деякі особливості.
Геном мітохондрій людини відносять до так називаного „ощадливого” типу. Основна інформація закодована у важкому ланцюзі (Н-ланцюг) ДНК (у ній локалізуються гени двох високомолекулярних рРНК, 14 тРНК і 12 генів, які кодують іРНК для білків внутрішньої мембрани мітохондрій). У легкому ланцюзі (L-ланцюг) ДНК зберігається інформація тільки про 8 тРНК і одну іРНК для єдиного білка малої субодиниці мітохондріальних рибосом.
У мітохондріях рослин кодується низькомолекулярна РНК рибосом. У мітохондріальних геномах хламідомонади й інфузорії тетрахімени зберігається інформація не про всі тРНК: частина тРНК, які працюють у мітохондріях, закодована в геномі ядра. А в ядерному геномі трипаносомід представлена інформація про всі тРНК мітохондріальні білоксинтезуючі системи. Істотні розходження спостерігаються між різними об'єктами й у кодуванні білків дихального ланцюга й АТФ-синтазного комплексу. Таким чином, на тлі відносної сталості інформаційної ємності мітохондріального генома виявляється і значна еволюційна пластичність цієї ознаки.
1.3 Функції мітохондрій
Різноманітні функції мітохондрій настільки тісно зв'язані з їхньою структурою, що їх зовсім неможливо вивчати окремо.
Ще перші цитологи, починаючи з Альтмана, припускали, що мітохондрії зв'язані з процесами окислювання в клітинах. У 1912 р. Бателі і Штерн, а потім у 1913 р. і Варбург встановили, що дихальні ферменти утримуються в нерозчинній фракції клітини. Про це відкриття не згадували до 1929 р., коли Кейліну вдалося виділити з м'язового волокна частки, які містять сукцинат- і цитохромоксидази. Це була перша поліферментна система, виділена з клітки, але її не вдалося ідентифікувати специфічними цитологічними методами. Класична робота Бенслі і Херра, які виділили гранулярну фракцію з клітин печінки і ідентифікували ці гранули з мітохондріями, проклала шлях сучасному вивченню ферментативних функцій мітохондрій. Було встановлено, що ізольовані мітохондрії активно окисляють глутамінову і янтарну кислоти і дають позитивну реакцію на цитохромоксидазу з реактивом нади. Було також виявлено, що при дегенерації мітохондрій, яка була викликана заморожуванням і наступним відтаванням клітини, в останньому знижується споживання кисню й активність сукцинатоксидази і цитохромоксидази. Нарешті, Хогебум, Клод і Хочкісс у 1946 р. довели, що обидва ці ферменти зв'язані з мітохондріальною фракцією клітини. З тих пір виділення мітохондрій стало широко розповсюдженою процедурою, і у вивченні функцій мітохондрій були досягнуті великі успіхи.
Ще з часів перших досліджень удалося установити, що мітохондрії складаються на 65-70% з білка і на 25-30% з ліпідів, головним чином фосфатидів (лецитину і кефаліну); холестерин та інші ліпіди присутні лише в незначній кількості. РНК незмінно складає близько 0,5% сухої ваги мітохондрій.
Існують два основних способи дослідження виділених мітохондрій: або їх вивчають у виді інтактних часток, або ці частки розділяють на усе більш і більш дрібні фрагменти, кожний з яких містить певні групи, які володіють ферментативною активністю. Обидва шляхи дослідження виявилися надзвичайно плідними і забезпечили одержання важливих даних про макромолекулярну і функціональну організацію мітохондрій.
Ферментна система мітохондрій. Ферментативні процеси мітохондрій вкрай складні; щоб їх охарактеризувати кількісно, достатньо сказати, що в реакції, які звичайно протікають у кожній мітохондрії, беруть більш 70 ферментів і коферментів, не рахуючи великого числа кофакторів, вітамінів і деяких металів.
Єдиною „сировиною”, яку потребують мітохондрії, є фосфат і аденозиндифосфат (АДФ), а кінцевим продуктом - АТФ, СО2 і Н2О. Три основні харчові речовини клітини (вуглеводи, жири і білки) в остаточному розщеплюються в цитоплазмі до двовуглецевих одиниць. Кожна така одиниця вступає в з'єднання з коферментом А, в результаті чого утворюється ацетилкофермент А. Коли останній попадає в мітохондрію, його ацетатна група вступає в цикл Кребса (його називають також циклом трикарбонових кислот або циклом лимонної кислоти) і після ряду складних реакцій за участю різних ферментів проходить декарбоксилювання, тобто вуглець, який входить до його складу виділяється у вигляді СО2. На різних етапах циклу пари електронів (або еквівалентне число атомів Н) під дією дегідрогеназ переносяться в дихальний ланцюг (ланцюг переносу електронів); зрештою вони з'єднуються з молекулярним киснем, утворюючи молекули води.
Дихальний ланцюг являє собою головну систему перетворення енергії в мітохондріях. Її компоненти не тільки функціонально, але і просторово тісно зв'язані зі структурою мітохондрій. До складу основних компонентів дихального ланцюга входять два флавопротеїдних фермента (сукцинат- і ДПН-дегідрогенази), чотири цитохроми, а також негемінове залізо, мідь і кофермент Q. У трьох точках цього ланцюга за рахунок енергії електронів з АДФ і фосфату утвориться АТФ. Тому говорять, що дихальний ланцюг сполучений з фосфорилюванням.
Локалізація ферментної системи мітохондрій. Один з методів вивчення локалізації ферментів у мітохондріях складається в подрібненні цих часток на фрагменти різного розміру, які мають різними властивостями. Якщо зважку мітохондрій піддати дії ультразвуку, то розчинні білки, які містяться в матриксі виділяються і в осадженій фракції залишаються частини мембран. Наприклад, у мітохондріях із клітин печінки утримується більше розчинних білків, чим у мітохондріях серцевого м'яза, які мають більш компактну структурою і велику кількістю крист.
До числа розчинних білків, які містяться в матриксі, відноситься більшість ферментів, які беруть участь у циклі Кребса (аконітаза, дегідрогеназа малеїнової кислоти, фумараза, дегідрогеназа ізолимоної кислоти, конденсуючий фермент, дегідрогенази піровиноградної і кетоглутарової кислот і т.д.) і жирних кислот (кротоназа, ацилдегідрогеназа). У матриксі мітохондрій містяться також різні нуклеотиди, нуклеотид-коферменти і неорганічні іони (К+, HPO4-, Mg++ Cl-- і SO4-). У нерозчинної, фракції, тобто в мембранах, знаходяться всі ферменти, які відносяться до дихального ланцюга, а також ферменти, які каталізують сполучений з нею процес окисного фосфорилювання.
Так як між числом крист і інтенсивністю реакцій дихального ланцюга існує пряма залежність, то було висунуто припущення, що в кристах і внутрішніх мембранах мітохондрій; міститься більшість ферментів, які беруть участь у цих реакціях.
До дуже важливого результату привели спектрофотометричні дослідження: вони дають підставу думати, що компоненти дихального ланцюга знаходяться в еквімолекулярних співвідношеннях, тобто на кожну молекулу цитохрому або цитохромоксидази припадає одна молекула сукцинатдегідрогенази або ДПН-дегідрогенази.
Деякі фрагменти мітохондрій, які утворюються під дією ультразвуку або отримані за допомогою інших методів роздроблення, ще зберігають здатність до окисного фосфорилювання, але при їх подальшому подрібнюванні отримують частки, які хоча і містять ферменти дихального ланцюга, однак не можуть забезпечити сполучення окислювання з синтезом АТФ.
Розрахунки показують, що 25-40% білків мембран складають ферменти дихального ланцюга, а 60-70% структурних білків не містять окислювально-відновних груп.
Ще однією стороною діяльності мітохондрій участь у специфічних синтезах, наприклад у синтезі стероїдних гормонів і окремих ліпідів. У мітохондріях можуть накопичуватися різні речовини і деякі іони, особливо іони Са2+. У мітохондріях ооцитів різних тварин утворюється скупчення жовтка, при цьому вони втрачають свою основну функцію. Мітохондрії, які відпрацювали можуть також накопичувати продукти екскреції. Нарешті, у деяких випадках (печінка, нирки) мітохондрії здатні акумулювати шкідливі речовини й отруту, які попадають в клітину, ізолюючи їх від основної цитоплазми і частково блокуючи шкідливу дію цих речовин.
1.4 Походження мітохондрій
На підставі даних світлової мікроскопії можна було припускати існування трьох можливих шляхів утворення мітохондрій: розподіл передіснуючих мітохондрій, біосинтез їх de novo і формування з немітохондріальних структур.
Спочатку вважали, що мітохондрії, подібно хромосомам, несуть генетичну інформацію і розмножуються діленням, тобто що нові мітохондрії утворяться в результаті ділення раніше існуючих мітохондрій і наступного росту продуктів розподілу.
Припущення про виникнення мітохондрій de novo засновано головним чином на даних дослідів з центрифугуванням яєць морського їжака. Було виявлено, що в деяких фракціях, які не мають мітохондрій (судячи з результатів фарбування різними методами), потім розвивався повний набір цих органоїдів. Ці дані були витлумачені в тім змісті, що мітохондрії утворяться не з передіснуючих елементів. Однак цілком можливо, що мітохондрії були присутні в досліджених фракціях, але просто не були виявлені методами, які застосовувалися.
Третя гіпотеза базується на даних електронної мікроскопії, які свідчать про наявність безперервного зв'язку між мембраною мітохондрій і плазматичною мембраною або ендоплазматичною сіткою і ядерною оболонкою. На цій підставі було висловлене припущення, що мітохондрії утворюються в результаті росту і переміщення або плазматичної мембрани, або мембран вакуолярної системи клітини.
Аналогічні дані були отримані на рецепторі інфрачервоних променів у деяких змій. У нервових закінченнях цього органа утримується щільна маса мітохондрій, а на невеликій відстані від них розташовані складки мембрани аксона; отже, можна припустити, що в механізмі утворення мітохондрій беруть участь як мембрана аксона, так і матрикс аксоплазми. Варто нагадати, що в бактерій мезосоми також утворюються з плазматичної мембрани.
Проблему походження мітохондрій ні в якій мірі не можна вважати вирішеною. Досліди, які були проведені з використанням міченого холіну на мутанті Neurospora crassa з недостатністю холіну, показали, що, очевидно, маса мітохондрій зростає за рахунок безперервного-додаткового включення нових молекул лецитину у вже існуючу структуру, і, таким чином, виключили можливість утворення мітохондрій de novo.
Факти на користь тієї точки зору, що додатковий ліпопротеїдний матеріал включається на якомусь низькому рівні структури. Відповідно до ряду сучасних біохімічних даних, напівперіод життя мітохондрій печінки складає приблизно 5-10 днів; отже, синтез мітохондрій у клітині може являти собою безперервний процес. Якщо це так, то варто було б з'ясувати, яким чином усі ферменти, які утримуються в мембранах і матриксі, організуються в єдину систему, яка обумовлює повну функціональну активність мітохондрії. У цьому відношенні цікавим прикладом можуть служити дріжджові клітини, які на відміну від клітин вищих організмів здатні існувати як у присутності, так і у відсутності кисню. В останньому випадку (анаеробіоз) у цих клітинах відсутні цитохроми b і а, і, отже, не можна говорити про дихальний ланцюга в буквальному значенні цього слова. Було також виявлено, що під час анаеробіозу в дріжджових клітинах відсутні істинні мітохондрії, а присутня тільки особлива система мембран, яка містить дві первинні дегідрогенази дихального ланцюга. Під дією кисню ці мембрани зливаються, утворюють складки і перетворюються в істинні мітохондрії, які містять цитохроми.
РОЗДІЛ 2. МЕТОДИ ДОСЛІДЖЕННЯ МІТОХОНДРІЙ
Значних успіхів було досягнуто у вивченні мітохондрій, при цьому значну роль зіграла електронна мікроскопія, яка дала можливість виявити їх специфічну ультраструктурну організацію, а також розробка методів біохімічного аналізу цих органоїдів після виділення.
Електронна мікроскопія. Взаємозв'язок довжини хвилі світла і межі дозволу зберігається для будь-якої форми випромінювання, як для світлових променів, так і для електронів. Однак в останньому випадку межа дозволу істотно нижче. Довжина хвилі електрона зменшується зі збільшенням його швидкості. В електронному мікроскопі з напругою 100000 В довжина хвилі електрона дорівнює 0.004 нм, а відповідно до теорії, дозвіл такого мікроскопа складає 0,002 нм.
Загальна схема просвічуючого електронного мікроскопа (ПЕМ). Джерело випромінювання - нитка катода, яка випускає електрони з вершини циліндричної колони висотою біля двох метрів. Оскільки при зіткненні з молекулами повітря електрони розсіюються, у колоні повинний бути створений вакуум. Електрони, які випромінюються катодною ниткою, прискорюються найближчим анодом і проникають через малюсінький отвір, формуючи електронний промінь, який проходить у нижню частину колони. Уздовж колони на деякій відстані розташовані кільцеві магніти, які фокусують електронний промінь, подібно скляним лінзам, які фокусують промінь світла у світловому мікроскопі. Зразок через повітряний шлюз поміщають у вакуум колони, на шляху електронного пучка. Частина електронів у момент проходження через зразок розсіюється відповідно до щільності речовини в даній ділянці, залишок електронів фокусується й утворить зображення на фотопластинці або на фосфоресцируючому екрані.
В електронному мікроскопі не можна спостерігати живі об'єкти. Тому тканини фіксують, зшиваючи клітини і клітинні структури глутаральдегідом, а потім обробляють осмієвою кислотою. Зразки збезводнюють, фіксують смолами і нарізають тонким скляним або алмазним ножем.
Тонкі зрізи практично є двовимірними зрізами тканини і не дозволяють судити про тривимірну структуру клітинних компонентів. Тривимірне зображення можна одержати після реконструкції сотень серійних зрізів. В даний час розроблені більш прямі методи одержання тривимірного зображення. Один з них складається у вивченні зразка під скануючим електронним мікроскопом. Для одержання зображення в просвітчастому електронному мікроскопі використовують електрони, які проходять через зразок, а в скануючому електронному мікроскопі використовуються електрони, які розсіюються або випромінюються поверхнею зразка. У даному випадку зразок повинен бути зафіксований, висушений і покритий тонкою плівкою важкого металу. Потім зразок сканується дуже вузьким пучком електронів. У такий спосіб відбувається формування єдиного, цільного і значно збільшеного зображення.
Метод скануючої електронної мікроскопії забезпечує значну глибину фокусування; більш того, оскільки масштаби розсіювання електронів визначаються кутом поверхні стосовно променя, на зображенні виникають світлі і темні ділянки, які чергуються та створюють враження тривимірності. Але цей метод застосовують тільки для вивчення поверхні і його дозвіл порівняно невеликий (близько 10 нм з ефективним збільшенням приблизно 20 тис. раз). Даний метод практично не застосовується для вивчення субклітинних органел і використовується винятково для вивчення цілих кліток і тканин.
Просвітлюючий електронний мікроскоп можна використовувати для вивчення поверхні зразка з дуже великим збільшенням, спостерігаючи окремі макромолекули. На висушений зразок напиляється тонка плівка важкого металу. Метал напиляється під певним кутом, так що відкладення напиленої плівки в деяких місцях товстіше, ніж в інших. Цей процес відомий як відтінення, при ньому виникає ефект тіні, який створює враження тривимірності зображення.
Приготовані в такий спосіб зразки можуть бути досить малі і тонкі, щоб електронний промінь проникав крізь них; наприклад, таким способом можна аналізувати індивідуальні молекули, віруси і стінки кліток. Що ж стосується більш товстих зразків, то тут після відтінення необхідно видалити органічний матеріал клітки, при цьому на поверхні зразка залишиться тільки тонкий металевий відбиток або репліка поверхні. Ця репліка потім підсилюється вуглецевою плівкою, після чого її можна помістити на сітку і вивчати в звичайному електронному мікроскопі.
У клітинній біології особливо успішно використовуються два методи, засновані на одержанні механічних реплік. Один з них - метод електронної мікроскопії «заморожування-сколювання» - дає можливість вивчати внутрішню будову клітинних мембран. Клітки заморожують при температурі рідкого азоту. Заморожений блок потім розколюють лезом ножа. Відкол часто проходить через гідрофобну середину подвійного шару ліпідів, оголюючи внутрішню поверхню клітинних мембран. Утворену поверхню відколу, яка утвориться відтіняють платиною, органічний матеріал видаляють і вивчають отримані репліки в електронному мікроскопі.
Метод «заморожування - травлення» використовується для вивчення зовнішньої поверхні кліток і мембран. У даному випадку клітки заморожують при дуже низькій температурі і замороженому блоці розколюють лезом ножа. Вміст льоду навколо кліток (і в меншому ступені усередині кліток) знижують сублімацією води у вакуумі при підвищенні температури (процес називають вакуумним сушінням). Ділянки клітини, піддані такому травленню, потім відтіняють для готування платинової репліки.
Використовуючи для контрастування відтінення солями важких металів, можна спостерігати в електронний мікроскоп ізольовані макромолекули, наприклад, ДНК або великі білки, а після негативного контрастування можна розгледіти навіть дрібні деталі. При готуванні зразків для негативного контрастування досліджувані молекули наносять на тонку плівку вуглецю (практично прозору для електронів), потім її змочують концентрованим розчином солей важких металів, наприклад, уранілацетата. Після висушування зразка тонка плівка солей важких металів рівномірно покриває вуглецеву підкладку, за винятком ділянок, зайнятих адсорбованими макромолекулами. Речовина макромолекул більш проникна для електронів у порівнянні з ближніми ділянками, покритими солями важких металів; за рахунок цього виникає звернене або негативне зображення молекули.
В даний час можна спостерігати з високим дозволом навіть внутрішні деталі тривимірних структур, таких, як віруси. Для цього використовують метод кріоелектроної мікроскопії, де дуже тонкий (приблизно 100 нм), швидко заморожений шар вологого зразка поміщають на мікроскопічні ґрати. За допомогою спеціального пристосування гідратований зразок утримують при - 160сС у вакуумі мікроскопа. Таким способом можна спостерігати матеріал практично безпосередньо: без фіксації, фарбування і сушіння.
При біохімічному аналізі використовують такі методи дослідження, як гібридизація, імуноцитохімія з застосуванням моноклінних антитіл на електронно-мікроскопічному рівні. За допомогою біохімічних методів вивчали АТФ-синтетазні комплекси внутрішньої мембрани.
Хоча дослідження мітохондрій у живій клітині пов'язано з відомими труднощами через те, що вони мають низький показник переломлення, їх усе-таки можна легко спостерігати в культурах тканини in vitro, зокрема під фазово-контрастним мікроскопом або в темному полі звичайного мікроскопа.
При фазово-контрастному та інтерференційному мікроскопуванні проходження світла через живу клітку фаза світлової хвилі міняється відповідно до коефіцієнта рефракції клітки: світло, яке проходить через відносно тонкі або відносно товсті ділянки клітки, затримується, і його фаза відповідно зрушується стосовно фази світла, яке проходить через відносно тонкі ділянки цитоплазми. Як у фазово-контрастному, так і в інтерференційному мікроскопі використовуються ефекти інтерференції, які виникають при рекомбінації двох наборів хвиль, що і створюють зображення клітинних структур. Обидва типи світлової мікроскопії широко використовуються для спостереження живих клітин.
Найпростіший спосіб розглянути деталі клітинної структури - спостерігати світло, яке розсіюється різними компонентами клітини. У темнопольному мікроскопі промені від освітлювача направляються збоку і при цьому в лінзи мікроскопа попадають тільки розсіяні промені. Відповідно клітина виглядає як освітлений об'єкт на темному полі. Одним з основних переваг фазово-контрастної, інтерференційної і темнопольной мікроскопії є можливість спостерігати рух клітин у процесі мітозу і міграції.
Високий контраст, який досягається за допомогою комп'ютерної інтерференційної мікроскопії, дозволяє спостерігати навіть дуже дрібні об'єкти.
За допомогою мікроманіпулятора було встановлено, що мітохондрії являють собою відносно стабільні утворення, які можна, неушкоджуючи, переміщати мікроголкою усередині клітини. Їх питома вага вище питомої ваги цитоплазми. Під час ультрацентрифугування живих клітин при 200000-400 000 g мітохондрії накопичуються у відцентрового полюса в неушкодженому виді.
Зміна об’єму і форми in vivo. Прижиттєві спостереження становлять особливий інтерес тоді, коли вони доповнюються цейтраферною мікрокінозйомкою. У культурі фібробластів можна спостерігати безперервні і часом ритмічні зміни об’єму, форми і розподілу мітохондрій. У русі мітохондрій розрізняють два основних типи: коливальні рухи і переміщення з однієї частини клітини в іншу, причому під час інтерфази вони значно більш активні, чим протягом мітозу. Іноді ці органоїди прикріплюються до оболонки ядра найближче до ядерця. Нитковидні мітохондрії можуть розпадатися на гранули, які здатні знову об’єднуватися в нитки. У деяких випадках рух мітохондрій зовсім пасивний й обумовлюється плином цитоплазми.
Різкі зміни об’єму і форми мітохондрій можуть викликатися хімічними, осмотичними і механо-хімічними реакціями. У живих клітинах спостерігаються цикли скорочень з малою амплітудою, які пов'язані з процесом окисного фосфорилювання. Відомо, що ціаністі сполуки, динітрофенол і інші інгібітори окислювання викликають набрякання, а надлишок АТФ - скорочення мітохондрій. Ці явища відбуваються і при зміні осмотичного тиску середовища. Неорганічний фосфат, відновлений глутатіон, Са2+ і жирні кислоти викликають набрякання, а дія АТФ запобігає цьому. Явище скорочення обумовлюється наявністю в мітохондріях скорочувального білка, аналогічного актоміозину м'язів. Більшої амплітуди змін об’єму можна досягти шляхом одночасного застосування декількох факторів (найбільш ефективними є Са2+ і тироксин) . Можливо, що набряканням мітохондрій, яке приводять до роз'єднання процесів окислювання і фосфорилювання, пояснюється фізіологічна дія цього гормону при гіперфункції щитовидної залози. Інші гормони, у тому числі гормон росту, окситоцин, вазопресин, інсулін і деякі кортикостероїди, також викликають набрякання мітохондрій. Крім того, це явище спостерігається і при патологічних станах організму, обумовлених, наприклад, впливом канцерогенів і токсинів.
Фіксація і фарбування. Вітальне і суправітальне дослідження мітохондрій значно полегшується при фарбуванні їх слабкими розчинами януса зеленого, у якому вони набувають зеленувато-синього кольору. Таке фарбування пояснюється присутністю в мітохондріях цитохромоксидазної системи, яка підтримує барвник в окисленій (зафарбованій) формі. У навколишній цитоплазмі барвник відновлюється до безбарвної лейкоформи.
Уже давно відомо, що мітохондрії досить лабільні і легко ушкоджуються під дією фіксаторів. Тому всі методи фіксації мітохондрій засновані на стабілізації їх ліпопротеїдної структури шляхом тривалої дії окисних агентів (оксид осмію(IV), хромова кислота і біхромат калію). Як барвник застосовують залізний гематоксилін або кислий фуксин Серед цитохімічних методів фарбування мітохондрій, слід виділити такі, як реакція наді на цитохромоксидазу або застосування різних сполук тетразолію, які утворють за участю дегідрогеназ мітохондрій нерозчинні хромогени (формазан). Подібні цитохімічні дослідження проводилися і з використанням електронного мікроскопа, причому для виявлення сукцинатдегідрогенази застосовувався телурит калію, а для виявлення сукцинат- і НАД-Н2-дегідрогеназ - нітросиній тетразолій і тетранітро-синий тетразолій. Виявилося, що локалізація продуктів реакції в мітохондріях тісно зв'язана з локалізацією крист.
РОЗДІЛ 3. ДОСЛІДЖЕННЯ МІТОХОНДРІЙ НА СУЧАСНОМУ ЕТАПІ
Таблиця 3.1
№ п/п
Рік видання
Назва та номер журналу
Автор та назва статті
Сторінка
1
1998
„Цитологія і генетика” № 2
И.В. Твердохлебов „Гетерогенность митохондриального аппарата миокарда и механизмы ее формирования в раннем онтогенезе крыс”
С.8-12
2
1999
„Цитологія і генетика” № 6
А.Ф. Попова „Ультраструктура митохондрий и интенсивность дыхания клеток Chlorella vulgaris Beijer. При различных сроках клиностатирования”
С.8-13
3
2001
„Цитологія і генетика” № 2
Д.О. Климчук ”Варіації ультраструктурної організації мітохондрій і клітинах гороху in vitro”.
С. 36-41
4
2002
„Цитологія і генетика” № 1
Б.А. Насибуллин ”Особенности реакции популяции митохондрий в нейронах сенсорной коры при длительном воздействии низкочастотной вибрации на организм крыс”.
С. 40-45
№ п/п
Рік видання
Назва та номер журналу
Автор та назва статті
Сторінка
5
2002
„Цитологія і генетика” № 5
А.Я. Литошенко ”Эволюция митохондрий”
С.49-57
6
2003
„Цитологія і генетика” № 5
Е.С. Сытник, А.Ф. Парий, И.К. Комарницкий, Ю.Ю. Глеба, Н.В, Кучук ”Анализ ядерного и митохондрияльного геномов у транспластомных растений Salpiglossis sinuata, полученных путем переноса трансформированных пластид от гибрида N. Tabacum (+S. Sinuate).
С.3-8
У 1999 році приділялась увага кількісним морфологічним характеристикам трьох типів мітохондрій у зрілих кардіоміоцитах крис. Встановлено існування двох режимів роботи мітохондріального апарату в залежності від локалізації та стану міофібрилярного апарату. Проведена оцінка кількісного відношення між типами органел на етапах онтогенезу. В цьому ж році був виділений коливальний характер рівня дихання культури та зміна ультраструктури мітохондрій. Відмічено два піка підйому інтенсивності дихання – на початковому етапі дії кліностатування і в кінці логарифмічної фази росту культури, які співпадають з періодами перерозподілу популяції. Підвищення інтенсивності дихання клітин та зміна ультраструктури направлені на стимуляцію метаболізму клітин в процесі адаптації до умов навколишнього середовища.
У 2001 році у частині казусних клітин гороху поряд з мітохондріями типової організації були виявлені гігантські органели. Їх оболонка як і у мітохондрій складається з двох біологічних мембран, з яких внутрішня утворює інвагінації. Специфічною особливістю структурної організації незвичайних мітохондрій є наявність включень у їх матриксі. Останні мають вигляд електронно щільного тяжу, що складається з фібрилоподібних утворень і простягається паралельно повздовжньої вісі органел.
У 2002 році в експерименті на 66 білих пацюках вивчена дія низькочастотної вібрації на популяцію мітохондрій нейронів сенсорної кори. Результати досліджень показали етапність реакції мітохондріальної популяції на дію низькочастотної вібрації; схожість структурних перебудов мітохондрій при дії вібрації та гіпоксії; перехід до одноманітності структурно-функціональних характеристик мітохондрій в популяції в динаміці низькочастотної дії.
Також проводились роботи щодо вивчення еволюції мітохондрій. Результати сіквенсу мітохондріальних геномів свідчать про монофілетичне походження мітохондрій від еубактеріального предка, який відноситься до підрозділу α-протеобактерій. Протягом останніх років були визначені повні послідовності великої кількості мітохондріальних та еубактеріальних геномів. Ці результати свідчать про те, що мітохондріальний геном еволюціонував від одного предка, загального для всіх існуючих на даний час еукаріотів, та що мітохондріальна та ядерна компоненти еукаріотичної клітини виникли одночасно.
В 2003 році на прикладі представника з родини Пасльонових Salpiglossis sinuata був запропонований новий підхід щодо створення пластомних трансформантів. В результаті експериментів були отримані морфологічно подібні з Salpiglossis sinuata рослини стійкі до стрептоміцину. Аналіз показав, що отримані рослини містять трансформований пластом Salpiglossis sinuata, а мітохондріальна ДНК отриманих рослин відрізняється від обох батьківських.
На основі проведеного аналізу можна сказати, що на сучасному етапі дослідження мітохондрій проводяться в багатьох вузьких, глибоко специфікованих напрямках, які стосуються впливу зовнішніх умов на будову і функції мітохондрій, особливостей роботи ферментного апарату, генетики і еволюції мітохондрій.
ВИСНОВКИ
Проведено огляд літературних джерел на основі яких з’ясовано особливості будови та функції мітохондрій, їхню роль в забезпечені клітин енергією.
Визначені особливості генетичної системи мітохондрій, її структура та особливості генетичного коду, визначено межі її автономності.
Розглянуто теорії походження мітохондрій та основні шляхи їх успадковування.
З’ясовані основні методи дослідження мітохондрій, і, як бачимо, вони представлені методами генетики, цитології, молекулярної біології і біохімії.
Проведено аналіз сучасних українських наукових досліджень, які проводились з мітохондріями.
СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
Албертс Б. и др. Молекулярная биология клетки. В 5-ти томах. М.: Мир, 1986 - 1987.
Заварзин А.А, Харазова А.Д., Молитвин М.Н. Биология клетки: Общая цитология: Учебное пособие для биол. спец. высших учебн.заведений. – Спб.:Из-во С. – Петербург. Ун-та, 1992. – 318 с.
Зенгбуш П. Молекулярная и клеточная биология. В 3-х т. М.: Мир, 1982. – 342 с.
Ленинджер А. Митохондрия. - М.: Мир, 1966. – 315 с.
Марченко А.І. Цитологія: Навчальний посібник для ден. Та заоч. Відділення природничо-географічного факультету / Ніжин, 2003. – 66 с.
Ченцов Ю.С. Общая цитология: [Учебное пособие для биолог. спец. вузов] – 2-е изд., М.: Из-во МГУ, 1984. – 350 с.
Э.де Робертис, В. Новинский, Ф. Саэс. Биология клетки. – М.: Мир, 1967. – 473 с.

Нравится материал? Поддержи автора!
Ещё документы из категории биология:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ