Дефенсины беспозвоночных животных
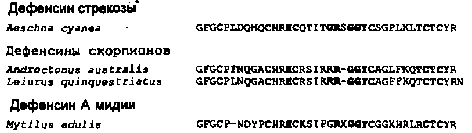
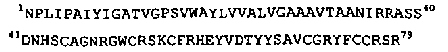


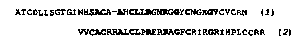
Реферат
по биологии
на тему:
"Дефенсины беспозвоночных животных"
2008
1. Дефенсины насекомых
Дефенсины насекомых были открыты независимо друг от друга двумя группами исследователей. Первоначально они были обнаружены в культуральной среде клеток NIH Sape-4, которые представляют собой поддерживаемую in vitro линию эмбриональных клеток серой мясной мухи Sarcophaga peregrina. Эти пептиды в литературе получили название сапецины. В 1989 г. французские исследователи выделили из гемолимфы личинки падальной мухи Phormia terranovae два антибактериальных пептида, получивших название формицин А и формицин В, которые имели частичное структурное сходство с дефенсином NP-1 из псевдоэозинофилов и альвеолярных макрофагов кролика.
Сравнение первичной структуры формицин а, выделенного из гемолимфы личинки падальной мухи, и дефенсина NP-1 из нейтрофилов кролика
Первичные структуры дефенсинов насекомых
Жирным шрифтам отмечены консервативные аминокислотные остатки. X - С-концевая часть роялизина: TSFK0LWDKYF.
В дальнейшем выяснилось, что эта гомология носит случайный характер и, как правило, не свойственна пептидам, принадлежащим к семейству дефенсинов насекомых, которых сейчас ухе известно более десятка. В частности, из представителей отряда двукрылых кроме формицинов и сапецинов были выделены и секвенированы дефенсины из мухи-пчеловидки Eristalis tenax, плодовой мушки Drosophila melanogaster, комара Aedes aegypti; из отряда жесткокрылых, или жуков, — из Zophobas atrarus, Tenebrio molitor, Allomyrina dichotoma; из отряда полужесткокрылых — из бескрылого красного клопа Pyrrhocoris apterus, зеленого древесного клопа Palomena prasina; из отряда перепончатокрылых — из медоносной пчелы Apis mellifera и шмеля; из отряда Odonata таким представителем является стрекоза Aeschia cyanea.
Первичная структура большинства представителей дефенсинов насекомых характеризуется заметной гомологией на всех участках молекул. Несколько особняком в сравниваемом ряду гомологичных пептидов стоит структура дефенсина из стрекозы Aeschna суа-пеа — представителя одного из наиболее эволюционно древних отрядов насекомых. Однако среди дефенсинов, выделенных из гемолимфы представителей «молодых» отрядов, также встречаются исключения, как в случае сапецина В из серой мясной мухи Sarcophaga peregrina. Дефенсины насекомых являются катионными молекулами, состоящими из 38—43 аминокислотных остатков. Исключение из этого правила составляют 2 пептида: — сапецин В и роялизин. Последний выделен как из маточного молочка пчел, так и гемолимфы взрослого животного. Для всех дефенсинов насекомых характерно присутствие 6 консервативно расположенных цистеиновых остатков, образующих стереотипно 3 внутримолекулярных дисульфидных мостика. Изучение надпервичной структуры формицина А методом ядерного магнитного резонанса выявило в его молекуле 3 различно организованных участка в пептидной цепи: гибкий и малоупорядоченный N-концевой, включающий аминокислотные остатки от 1-го до 13-го, центральный, представленный амфипатической а-спиралью, и третий — С-концевой, являющийся р-слоем, сформированным 2 антипараллельными Р-тяжами, которые связаны Р-изгибом. Дисульфидные мостики являются фактором, стабилизирующим вторичные структуры пептидов и поддерживающим их третичную структуру, которая необходима для максимальной реализации антимикробных свойств дефенсиновой молекулы. На основании сравнения первичных структур дефенсинов насекомых можно допустить, что для большинства из них характерна подобная надпервичная структура, подтвержденная экспериментально, однако, только в случае сапецинов. Она отлична от вторичной структуры а- и р-дефенсинов, которая представлена 3 антипараллельными р-тяжами. Отличная от а- и Р-дефенсинов аранжировка дисульфидных связей дефенсинов насекомых наряду с особенностями их первичной и вторичной структур определяет различия третичных структур рассматриваемых пептидов. У дефенсинов насекомых она ближе к трехмерной структуре пептидного нейротоксина из яда скорпиона — харибдотоксина, который является ингибитором К+-каналов. В связи с этим, по-видимому, не случайным является факт обнаружения у сапецина В способности ингибировать К+-каналы в клетках Пуркинье из мозжечка крысы.
Различия в структурах дефенсинов насекомых, с одной стороны, и а- и Р-дефенсинов — с другой, ответственны, по всей видимости, за особенности их антимикробной активности. Так, если дефенсины млекопитающих и птиц являются универсальными антибиотическими пептидами, инактивирующими грамположительные и грамотрицательные бактерии, низшие фибы и оболочечные вирусы, то этого нельзя сказать о дефенсинах насекомых, последние действуют преимущественно только на грамположительные бактерии. Необходимы на 1—2 порядка большие концентрации для проявления антимикробной активности дефенсинов насекомых в отношении грамотрицательных бактерий и еще более значительные — в случае воздействия на грибки. Ограниченная функциональная активность дефенсинов насекомых в отношении отдельных групп микробов компенсируется на уровне организма продукцией других антибиотических пептидов, таких как цекропины, апидацины, дрозоцин, метальниковины, которые эффективно инактивируют грамотрицательные бактерии, и дрозомицин, направленно воздействующий на грибки.
Изучение механизмов антимикробного действия дефенсинов насекомых выявило в принципе все те же закономерности проявления их активности, которые были установлены для а- и р-дефенсинов. Основной мишенью их поражающего действия на бактерии является цитоплазматическая мембрана. Перфорируя плазмалемму бактерий, дефенсины насекомых нарушают структурную целостность клетки, ее ионный го-меостаз, вызывают частичную деполяризацию мембраны и уменьшение концентрации цитоплазматического аденозинтрифосфата вследствие ингибирования окислительного фосфорилирования. Остается, однако, не совсем ясным вопрос, почему при столь общих проявлениях воздействия на цитоплазматическую мембрану дефенсины насекомых по сравнению с дефенсинами млекопитающих и птиц характеризуются столь слабой антимикробной активностью в отношении грамотрицательных бактерий. Может быть, фактором, ограничивающим антимикробную активность дефенсинов насекомых в отношении фамотрицательных бактерий, является наружная мембрана последних, которая слабо проницаема именно для рассматриваемого типа пептидных молекул.
У большинства изученных видов насекомых содержание антимикробных пептидов в гемолимфе находится на фоновом уровне. Это касается и присутствия таких пептидов, как цекропины и дефенсины насекомых. Синтез дефенсинов у насекомых, как и большинства других антибиотических пептидов, носит преимущественно индуцибельный характер. Попадание бактерий и грибов, а также липополисахаридов и продуктов их метаболизма во внутреннюю среду животных инициирует защитный процесс продукции жировым телом насекомых широкого спектра антимикробных пептидов, секретируемых в гемолимфу. Меньший вклад в биосинтез этих пептидов вносят гемоциты и другие клеточно-тканевые структуры организма. Антимикробные пептиды продуцируются также эпидермальными клетками в ответ на повреждение или инфицирование кутикулы насекомых. В этом наблюдается существенное сходство данного процесса с биосинтезом и продукцией Р-дефенсинов эпителием трахеи крупного рогатого скота. Более того, существует удивительное совпадение ряда молекулярно-генетических процессов в эпителиях у насекомого и млекопитающего, выражающееся в вовлечении в индуктивный синтез пептидов сходных генетических структур и, в частности, так называемых кр-подобных промоторных участков генов, которые ответственны за синтез антибиотических пептидов у столь эволюционно удаленных видов животных. Необходимо, однако, отметить, что если для насекомых подобный индуцибельный характер биосинтеза антибиотических пептидов является правилом, то для млекопитающих феномен индуцибельного синтеза р-дефенсинов в эпителии трахеи и кератиноцитах кожи человека — это пока, скорее всего, исключение. Для высших животных свойствен конститутивный синтез лейкоцитарных и эпителиальных антимикробных пептидов и белков только на определенной стадии их клеточного развития, с последующей упаковкой этих защитных соединений в гранулы на уровне аппарата Гольджи и сохранения их в соответствующих клетках в преформированном, уже, как правило, готовом для функционирования состоянии. Повреждения, инициирующие фагоцитоз или воспалительную реакцию, запускают процесс мобилизации депонированных в виде предшественников антибиотических пептидов и белков.
Контроль активности иммунных генов насекомых сходен с таковым генов иммуноглобулинов и острофазовых белков позвоночных. Они имеют в промоторной области генов энхансерные последовательности, получившие наименование кВ-мотивов, поскольку впервые были обнаружены в структуре генов, ответственных за синтез к-цепи иммуноглобулинов млекопитающих. кВ-Мотив является элементом гена, с которым избирательно связывается регуляторный транскрипционный фактор NFkB, обычно находящийся в цитоплазме в связанном с ингибитором состоянии. Воздействия на организм, запускающие иммунный ответ или воспалительный процесс, приводят к распаду функционально неактивного комплекса NFkbAkB, освобождению NFkB и его транслокации в ядро, где он взаимодействует с соответствующей промоторной областью генов острофазовых белков и иммуноглобулинов, активируя транскрипционный процесс. Для дефенсинов млекопитающих подобный механизм регуляции активности их генов описан пока только для де-фенсина трахеи крупного рогатого скота, в то время как для процессов регуляции биосинтеза миелоидных и энтеральных дефенсинов свойственны другие закономерности.
В настоящее время описана структура нескольких генов, ответственных за кодирование индуцибельных антибактериальных пептидов у Drosophila melanogaster, Hyalophora cecropia и Sarcophaga peregrina. Как правило, это — «молчащие» гены в условиях, когда отсутствуют клетки и вещества, инициирующие иммунный ответ. Активация таких генов при повреждении кутикулы или инфицировании насекомых происходит в течение получаса. Пик транскрипции наблюдается в течение 12—48 ч и зависит от характера и природы факторов, инициирующих иммунную реакцию. Некоторые из этих генов содержат небольшие интроны, хотя большинство генов не включают в свою структуру таковых. Как правило, антибиотические пептиды синтезируются в форме больших молекул, состоящих из 3 частей: сигнальной, прочасти и конечного зрелого пептида.
Проксимальная область данных генов содержит несколько последовательностей, имеющих структурную гомологию с цисрегуляторными элементами промоторов генов, ответственных за синтез острофазовых белков у млекопитающих. Чаще всего в таких структурах идентифицируется кВ-родственный элемент. Его существование у насекомых доказано и генетическими методами. В качестве дополнительных регуляторных элементов проксимальной области промотора выступают последовательности ДНК, гомологичные элементам млекопитающих, ответственных за реагирование к IL-6 и у-интерферону. Из клеток Hyalophora cecropia были выделены белки, относящиеся к Rel/NF-кВ-семейству, которые избирательно связываются с соответствующей областью промотора при инициации иммунного ответа у насекомых. Это, в частности, так называемый фактор, ответственный за иммунную реакцию у цекропии {Cecropia immune-responsiveness factor; Sun, Faye, 1992a, 1992b). У дрозофилы он носит название Dif (dorsal-related immunity factor) и связывается специфически с промоторами генов, ответственных за синтез антибиотических пептидов. В условиях инициации иммунного ответа Dif переносится из цитоплазмы в ядро и запускает процессы активации соответствующих генов. Как правило, эти процессы протекают в клетках жирового тела, эпителия кутикулы и гемоцитах насекомых {Hyalophora cecropia, Drosophila melanogaster, Bombyx tnori).
Выявленное сходство в механизмах регуляции генов насекомых, ответственных за синтез индуцибельных антибиотических пептидов, и генов млекопитающих, определяющих структуру острофазовых белков и Р-дефенсина эпителия трахеи, свидетельствует о существовании ряда единых принципов молекулярной организации и регуляции генов в процессе эволюции животного мира.
2. Дефенсины скорпионов
Скорпионы принадлежат к тому же типу членистоногих, что и класс насекомых. Они по одной из классификаций составляют подкласс Scorpiones в составе класса Arach-nida, входящего в подтип Chelicerata. Поэтому с позиции сравнительной биохимии представляет интерес оценка структурной гомологии антибиотических пептидов из гемолимфы скорпионов и о эволюционно близких им насекомых. В настоящее время выделены и секвенированы дефенсины из гемолимфы 2 видов скорпионов: Leiurus quinquestriatus и Androctonus australis, которые имеют максимальную степень структурной гомологии с дефенсинами из гемолимфы стрекозы Aeschna суапеа, представителя наиболее архаичного, а следовательно, и наиболее близкого скорпионам по эволюционному происхождению отряда насекомых.
3. Дефенсины мечехвостов
Мечехвосты относятся к подклассу Xiphosura класса меростомовых подтипа хелицеровые, входящего в тип членистоногих. Это одна из древнейших, сохранившихся до наших дней группа членистоногих животных, а потому естественным представляется интерес к факту обнаружения в гемоцитах японского подковообразного краба пептидов, подобных дефенсинам:
Сравнение первичной структуры дефенсинов, относящихся к классу насекомых, классу паукообразных и классу пластинчатожаберных, или двустворчатых
Этот пептид получил название «большой дефенсин», так как он состоит из 79 аминокислотных остатков, 37 из которых в С-концевой области имеет сходство по последовательности и наличию 3 дисульфидных мостиков с дефенсином RtNP-2 из нейтрофилов крысы. Молекула пептида состоит из 2 больших функциональных блоков: N-концевого слабоосновного гидрофобного из 42 аминокислот и С-концевого, имеющего черты структурного сходства с миелоидным дефенсином крысы по первичной структуре, а по аранжировке дисульфидных мостиков аналогичного Р-дефенсинам из нейтрофилов коров. Таким образом, рассматриваемый дефенсин по своим структурно-функциональным свойствам выпадает из круга семейства дефенсинов насекомых, характерных для ряда видов насекомых, скорпионов и моллюсков.
4. Дефенсины моллюсков
Моллюски являются вторым крупным типом первичноротых беспозвоночных, отдельные представители которого достигли в процессе эволюции высокого уровня морфофункционального развития. Более 545 млн лет тому назад, в геологический период кембрия, 2 основных, наиболее развитых типа беспозвоночных — членистоногие и моллюски — разошлись в своем магистральном эволюционном развитии. Общие морфогенетические корни рассматриваемых типов предполагают наличие у ряда их представителей сходных молекул, обеспечивающих процессы развития и иммунитета. В совместном российско-французском проекте были выделены и изучены антимикробные пептиды из гемолимфы мидии Mytilus edulis, представителя класса пластинчатожаберных, или двустворчатых моллюсков. Среди нескольких групп пептидов были охарактеризованы и представители дефенсинов. По своей первичной структуре они оказались молекулами, максимально гомологичными дефенсинам стрекозы Aeschna суапеа и 2 видов скорпионов — Leiurus quenquestriatis и Androctonus australis.
По заключению исследователей, структурная гомология дефенсинов, выделенных из гемолимфы представителей двух типов, может свидетельствовать об их происхождении от общего молекулярно-генетического предшественника. Интересно, что у мидии, как и у скорпионов, дефенсины присутствуют в гемолимфе конститутивно, независимо от факторов микробной природы и повреждений, ведущих к воспалительной реакции и индуктивно повышающих продукцию дефенсинов в организме насекомых. В связи с этим фактом представляет интерес выяснение различий в молекулярно-генетических основах биосинтеза дефенсинов у насекомых, с одной стороны, и скорпионов и моллюсков — с другой.

Нравится материал? Поддержи автора!
Ещё документы из категории биология:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ