Физиология рыб
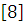
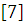
 Калининградский Государственный Технический Университет
Калининградский Государственный Технический Университет
Контрольная работа
По дисциплине:
«Физиология рыб»
Выполнил:
Плотников Виктор Александрович
Студент группы: 07-ЗВА
Шифр- 3188
Калининград 2010
Содержание
Вопрос №24. Строение и функция обонятельного анализатора. Вкусовая рецепция
Вопрос №13. Что такое рецепторы; афферентные, эфферентные и вставочные нейроны?
Вопрос №31. Каковы функции печени? Опишите состав желчи и её роль в пищеварении
Вопрос №3. Что такое раздражимость, возбудимость, возбуждение? Каковы признаки возбуждения? Что такое раздражители? Какие бывают раздражители? Что такое порог раздражения? Что такое торможение?
Список используемой литературы
Вопрос №24. Строение и функция обонятельного анализатора. Вкусовая рецепция
Восприятие животными химических веществ с помощью рецепторов – одна из форм реакции организмов на воздействие внешней среды. У водных животных специализированные рецепторы контактируют с веществами, находящимися в растворенном состоянии, поэтому характерное для наземных животных четкое разделение на рецепторы обоняния, воспринимающие летучие вещества, и рецепторы вкуса, воспринимающие вещества, находящиеся в твердом и жидком состоянии, у водных животных не проявляется. Однако морфологически и функционально органы обоняния у рыб достаточно хорошо обособлены. Ряд исследователей, основываясь на отсутствии специфичности в функционировании, локализации и связи с нервными центрами, объединяют вкус и общее химическое чувство понятием «химический анализатор», или «необонятельная хеморецепция».
Рецепторы обоняния у рыб, как и у прочих позвоночных, парные и расположены в передней части головы. Лишь у круглоротых они непарные. Обонятельные рецепторы находятся в слепом углублении – ноздре, дно которой выстлано обонятельным эпителием, располагающимся на поверхности складок. Складки, расходясь радиально от центра, образуют обонятельную розетку.
У разных рыб обонятельные клетки расположены на складках по-разному: сплошным слоем, разреженно, на гребнях или в углублении. Ток воды, несущий молекулы пахучих веществ, попадает к рецептору через переднее отверстие, часто отделенное от выходного заднего лишь складкой кожи. Однако у некоторых рыб входное и выходное отверстия заметно разделены и далеко отстоят друг от друга. Передние (входные) отверстия у ряда рыб (угорь, налим) находятся близко к концу рыла и снабжены кожными трубочками. Считается, что этот признак свидетельствует о значительной роли обоняния в поисках пищевых объектов. Движение воды в обонятельной ямке может создаться либо движением ресничек на поверхности выстилки, либо сокращением и расслаблением стенок специальных полостей – ампул, либо в результате движения самой рыбы.
Рецепторные обонятельные клетки, имеющие биполярную форму, относятся к категории первичных рецепторов, т.е. сами генерируют импульсы, содержащие информацию о раздражителе, и передают их по отросткам к нервным центрам. Периферический отросток обонятельных клеток направляется к поверхности рецепторного слоя и заканчивается расширением – булавой, на апикальном конце которой имеется пучок волосков, или микроворсинок. Волоски пронизывают слой слизи на поверхности эпителия и способны к движениям. Обонятельные клетки окружены опорными клетками, которые содержат овальное ядро и многочисленные гранулы разного размера. Здесь же расположены базальные клетки, не содержащие секреторных гранул. Центральные отростки рецепторных клеток, не имеющие миелиновой оболочки, пройдя базальную мембрану эпителия, образуют пучки до нескольких сотен волокон, окруженные мезаксоном шванновской клетки, причем тело одной клетки может охватывать много пучков. Пучки сливаются в стволики, образующие обонятельный нерв, соединяющийся с обонятельной луковицей.
Рис.1 Строение обонятельной выстилки: 1 – ресничная клетка; 2 – грушевидная секреторная клетка; 3 – обонятельные луковицы; 4 – опорные клетки; 5 – базальные клетки; 6 – бокаловидная секреторная клетка.
Обоняние как способ получения информации об окружающем мире имеет для рыб весьма существенное значение. По степени развития обоняния рыб, как и других животных, обычно делят на макросматиков и микросматиков. У макросматиков органы обоняния способны воспринимать большое количество различных запахов, т.е. обоняние используется ими в более разнообразных ситуациях. Типичным представителем является обыкновенный угорь. Микросматики воспринимают обычно небольшое количество запахов - в основном особей своего вида и половых партнеров. Типичные представители: щука, трехиглая колюшка.
Чувствительность рыб к запахам очень велика. Для восприятия запаха иногда, видимо, достаточно попадания на обонятельный рецептор нескольких молекул вещества.
Обоняние может играть руководящую роль в поисках пищи, особенно у ночных и сумеречных хищников, таких, как угорь. С помощью обоняния рыбы могут воспринимать партнеров по стае, находить особей другого пола в период размножения. Рыбы одного вида способны воспринимать химические соединения, выделяемые кожей других рыб при ранении. Эти «вещества тревоги» также воспринимаются органами обоняния и играют роль сигнала опасности для других особей в стае.
Работа над ошибками по замечаниям на вопрос №24
Строение обонятельной выстилки у всех позвоночных сходно (Рис.1), что свидетельствует о сходстве механизма обонятельной рецепции. Однако сам этот механизм еще не вполне ясен. Существует ряд гипотез на этот счет. Одна из них связывает возможности распознавания запахов, т.е. молекул пахучих веществ, с избирательной специфичностью индивидуальных рецепторов к запахам. Такова стереохимическая гипотеза Эймура, согласно которой имеется семь типов активных мест на обонятельных клетках, которые соответствуют семи основным запахам, а молекулы веществ, обладающих сходными запахами, имеют одинаковую форму активных частей, которые подходят к активным точкам рецептора, как ключ к замку. Другие гипотезы связывают возможность различения запахов с различиями в распределении веществ, адсорбируемых слизью обонятельной выстилки, по ее поверхности. Ряд исследователей полагают, что распознавание запахов обеспечивают два механизма, дополняя друг друга. Ведущая роль в обонятельной рецепции принадлежит, видимо, волоскам и булаве обонятельной клетки, которые обеспечивают специфическое взаимодействие молекул одорантов с мембраной клетки и перевод эффекта взаимодействия в форму электрического потенциала.
Аксоны обонятельных рецепторных клеток образуют обонятельный нерв, входящий в обонятельную луковицу, являющуюся первичным центром обонятельного рецептора. Обонятельная луковица относится, как считает А.А.Заварзин, к экранным структурам. Для нее характерно расположение элементов в виде последовательных слоев, причем нервные элементы связаны между собой не только в пределах слоя, но и между слоями. Таких слоев выделяют обычно три: слой обонятельных клубочков с межклубочковыми клетками; слой вторичных нейронов с митральными и кисточковыми клетками; гранулярный слой. Информацию в высшие обонятельные центры у рыб передают вторичные нейроны и клетки гранулярного слоя. Наружная часть обонятельной луковицы состоит из волокон обонятельного нерва, контакт которых с дендритами вторичных нейронов происходит в обонятельных клубочках, где наблюдается ветвление тех и других окончаний. Обонятельные луковицы рыб анатомически хорошо обособлены и бывают двух типов: сидячие и стебельчатые.
Необонятельная хеморецепция у рыб осуществляется вкусовыми почками и свободными окончаниями блуждающего, тройничного и некоторых спинномозговых нервов. Структура вкусовых почек сходна у всех классов позвоночных. У рыб они обычно имеют овальную форму и состоят из 30 – 50 вытянутых клеток, апикальные концы которых образуют канал. К основанию клеток подходят окончания нерва. Это типичные вторичные рецепторы. Они располагаются в ротовой полости, на губах, жабрах, в глотке, на коже головы и туловища, на усиках и плавниках. Их количество варьирует от 50 до сотен тысяч и зависит, как и их расположение, в большей мере от экологии, нежели от вида. Размеры, количество и распределение вкусовых почек характеризует степень развития вкусовой рецепции конкретного вида рыб.
Вкусовые почки передней части рта и кожи иннервируются волокнами возвратной ветви лицевого нерва, а слизистой рта и жабр – волокнами языкоглоточного и блуждающего нервов. В иннервации вкусовых рецепторов участвуют также тройничный и спинальные нервы.
Химические соединения воспринимаются также нервными окончаниями и сенсорными аппаратами в виде цилиндров, каналов, веретен, кустиков. Установлено, что передняя часть боковой линии и ампулы Лоренцини хрящевых рыб являются рецепторами, воспринимающими изменения солености воды. Центры вкусовой и общей химической чувствительности находятся в продолговатом мозге, в вагусной доле или обособленной доле лицевого нерва. Эти центры имеют связи с ядрами таламуса и гипоталамуса, а также среднего мозга. Механизм вкусовой рецепции в общих чертах выяснен. Четыре основных вкусовых ощущения – сладкое, горькое, кислое и соленое – воспринимаются благодаря взаимодействию молекул вкусовых веществ с четырьмя типами белков. Комбинации этих типов и создают конкретные вкусовые ощущения. У большинства рыб вкус играет роль контактной рецепции, поскольку пороги вкусовой чувствительности сравнительно высоки. Но у некоторых рыб вкус может приобретать функции дистанционного рецептора.
Вопрос №13. Что такое рецепторы; афферентные, эфферентные и вставочные нейроны?
Жизнь рыбы становится возможной благодаря наличию адекватных реакций её организма на изменения, происходящие во внешней среде и в самом организме. Получение информации о физических и химических характеристиках среды называется рецепцией. Структуры организма, воспринимающие эту информацию, называются рецепторами, т.е. приемниками. Рыба реагирует на световое излучение и внешний вид объектов материального мира, т.е. обладает зрением; реагирует на температуру окружающей среды, т.е. обладает терморецепцией; на её поведение влияют многие растворенные в воде вещества, т.е. она обладает химической рецепцией – вкусом и обонянием; воспринимает упругие колебания среды – слышит. Рыба способна к проприорецепции – анализу своей позы и положения тела, кроме того, она способна реагировать на ускорения, вызываемые смещением тела. Легко обнаруживается тактильная рецепция рыб – реакция на прикосновения, удар, давление. На рыб оказывает влияние электрический ток, а также имеются указания, что в некоторых условиях она может реагировать на магнитное поле. На радиоактивное излучение сублетальной интенсивности рыбы, как и другие живые существа, не реагируют.
В основе деятельности центральной нервной системы лежит рефлекс. Рефлекс – это реакция организма на раздражение рецепторов различными изменениями внутренней или внешней среды при участии центральной нервной системы, в которой происходит разложение попадающих на рецепторы раздражений, а затем их объединение и синтез.
Существует несколько квалификаций рефлексов. И.П. Павлов разделил все рефлексы по их происхождению на две большие группы: рефлексы врожденные, безусловные, которые передаются животному по наследству и которые сохраняются у него в течение всей жизни (видовые рефлексы); рефлексы условные, которые приобретаются в течении жизни, и если их не подкреплять, они угасают.
В зависимости от расположения раздражаемого рецептора выделяют экстерорецептивные рефлексы, возникающие при раздражении наружной поверхности тела (кожи, слизистых), интерорецептивные, возникающие при раздражении рецепторов внутренних органов, и проприорецептивные – при раздражении рецепторов скелетных мышц, суставов, связок.
Рефлексы делятся также в зависимости от того, какой отдел необходим для их вызывания: спинальные (спинномозговые) – для осуществления этих рефлексов необходимы центры спинного мозга; бульбарные – необходимые центры продолговатого мозга; мезенцефальные – центры среднего мозга; диэнцефальные – центры промежуточного мозга. Иногда разделяют рефлексы по тому органу, который участвует в ответной реакции. Если это будет мышца, то рефлексы называются моторными, двигательными, если железа – секреторными, если сосуд – сосудодвигательными.
Рефлекторная дуга представляет собой совокупность различных образований нервной системы, участвующих в выполнении рефлекторных реакций. (Рис.2). В состав рефлекторной дуги входят следующие образования.
1. Рецепторы – нервные окончания, в которых энергия внешнего раздражения превращается в нервные импульсы.
2. Афферентные (чувствительные) нервные волокна – отростки биполярных клеток спинальных ганглиев, по которым возбуждающие нервные импульсы идут в центральную нервную систему.
3. Нервный центр – участок центральной нервной системы, где афферентные импульсы подвергаются обработке – суммированию, трансформации, усилению или торможению, а также переключению на соответствующий эфферентный нейрон.
4. Эфферентные (центробежные) нервные волокна, по которым нервные импульсы передаются в органы-исполнители к мышечным, пигментным или секретирующим клеткам.
К рефлекторной дуге также относятся афферентные нервные пути, передающие в центральную нервную систему информацию от рецепторов функционирующих органов о характере деятельности, что позволяет регулировать на основе механизма обратной связи осуществление рефлекторной деятельности.
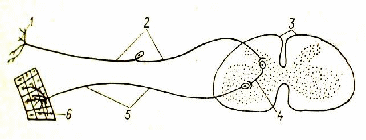
Рис.2. Рефлекторная дуга:
1 – кожный рецептор; 2 – афферентное волокно; 3 – спинной мозг; 4 – вставочный нейрон; 5 – эфферентное волокно; 6 – мышца.
Эфферентные пути вегетативной и соматической нервной систем разные. Хотя в пределах спинномозговых нервов все эфферентные импульсы выходят по вентральному корешку, вскоре этот корешок у рыб делится на три веточки. По двум веточкам импульсы идут к мышцам, а по третьей – висцеральной – к внутренним органам. В то же время у вегетативной и соматической нервной систем имеются и общие черты. Афферентные пути у этих двух систем общие. Вегетативная нервная система не имеет собственных афферентных путей. Обе системы регулируются центрами головного мозга. В целостной реакции организма при выполнении поведенческих условнорефлекторных реакций участвуют как соматическая, так и вегетативная нервная система. Последняя способствует лучшему кровоснабжению органов и улучшению в них обмена веществ.
Работа над ошибками по замечаниям на вопрос №13
Афферентные (чувствительные, центростремительные) нейроны являются отростками биполярных клеток, они чувствительные нейроны, несущие информацию в центральную нервную систему (спинной, головной мозг).
Эфферентные (центробежные, двигательные) нейроны несут информацию из центральной нервной системы к периферии (к мышцам и рабочим органам).
Вставочные нейроны (контактные, ассоциативные), находятся в пределах центральной нервной системы, передают информацию от афферентных нейронов к эфферентным.
Наглядный пример Рис.2.
Вопрос №31. Каковы функции печени? Опишите состав желчи и её роль в пищеварении
В начальную часть кишечника впадают протоки двух пищеварительных желез: печени и поджелудочной железы.
В печени происходит обезвреживание ядовитых веществ, поступающих из кишечника, вырабатывается желчь, эмульгирующая жиры и усиливающая перистальтику кишечника, осуществляется синтез белков и углеводов, накапливаются гликоген, жир, витамины (особенно у акул и тресковых).
У хрящевых рыб печень трёхдольчатая. Масса её достигает 10-20 % массы тела. У многих карповых печень также состоит из трех долей, но у сазана (карпа) их две, а у щуки и окуня одна.
Двухдольчатую печень имеют многие костистые рыбы.
У многих рыб, в частности у карповых, печень включает ткань поджелудочной железы и называется гепатопанкреасом.
Большую роль в пищеварении играют печень и её секрет – желчь. Желчь вырабатывается клетками печени и по дренажной системе желчных протоков поступает в желчный пузырь. В пузыре желчь сгущается до содержания сухого вещества приблизительно 13 %, более половины которого составляют желчные кислоты. Зеленоватый или коричневатый цвет желчи обусловлен присутствием желчных пигментов, представляющих собой продукты деградации гематина. Желчь содержит значительное количество минеральных катионов и анионов – кальция, магния, сульфата, карбоната. В ней содержатся слизистые мукоиды и полярные липоиды, играющие роль смазки для пищевых частиц. В желчи обнаруживается амилазная, липазная и протеазная активность.
Среди веществ желчи основную роль играют поверхностно-активные вещества – желчные кислоты. Они же обусловливают хорошо всем известную горечь желчи. Понижать поверхностное натяжение на границе водной среды и жировых капель способны также желчные пигменты. Поверхностно-активные вещества способствуют распаду жировых капель и стабилизации жировой эмульсии, что увеличивает поверхность пищевого жира, доступную липолитическим ферментам. Кроме того, сверхтонкая эмульсия жира может усваиваться без окончательного гидролиза. Желчные кислоты могут соединяться с высшими жирными кислотами, делая их растворимыми в воде, что улучшает их усвоение. Желчные кислоты, всосавшиеся вместе с жиром, снова попадают в печень и используются повторно.
Желчный пузырь представляет собой как бы боковой карман главного желчного протока. Когда желчный проток перекрыт путем сужения просвета ниже желчного пузыря, желчь накапливается в пузыре, гладкая мускулатура которого в это время расслаблена и сам он растянут. Такое состояние пузыря наблюдается в отсутствие пищеварения. У зимующей молоди карпа желчный пузырь раздут и достигает размеров ягоды смородины. Специфические гормональные воздействия, связанные с питанием и пищеварением, заставляют расслабляться сфинктер желчного протока и напрягаться стенки пузыря. В результате этого желчь попадает в просвет кишечника. Устье желчного протока в кишечнике рыбы определяется по желто-коричневому пятну на отмытой слизистой.
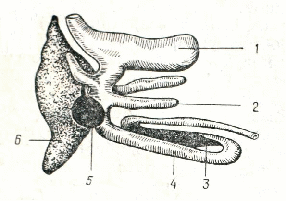
Рис.3 Часть пищеварительного тракта окуня.
1 – желудок; 2 – пилорические придатки; 3 – селезенка; 4 – кишечник; 5 – желчный пузырь; 6 – печень.
Вопрос №3. Что такое раздражимость, возбудимость, возбуждение? Каковы признаки возбуждения? Что такое раздражители? Какие бывают раздражители? Что такое порог раздражения? Что такое торможение?
Нервная система объединяет и согласует деятельность всех систем организма, в результате чего организм способен правильно реагировать на изменение внешней и внутренней среды. Нервная система состоит из центральной и периферической. В центральной нервной системе происходит переработка поступающей информации и принимаются целесообразные решения, а периферическая нервная система передает информацию и точно адресованные команды.
При возбуждении нервного волокна происходит деполяризация, т.е. уменьшение, или инверсия, поляризации мембраны. Нервное возбуждение распространяется по нервам в виде нервных импульсов (спайков), которые характеризуются быстронарастающим и спадающим возбуждением, распространяющимся по нейрону и сопровождающимся изменением физико-химических свойств и электрического потенциала возбужденного участка. Этот процесс является самораспространяющимся от источника возбуждения и происходит он без затухания. Повреждение нервного волокна нарушает проведение импульса. Возбуждение, передающееся по какому-либо нервному волокну, не передается на соседние нервные волокна этого же нервного ствола. Возбуждение по нервному стволу может одновременно передаваться в двух направлениях – центростремительном и центробежном.
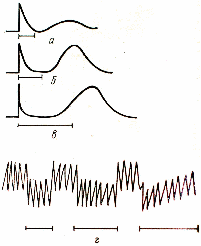
Раздражение вагуса ведет к уменьшению сердечного тонуса. Частота сердцебиений не уменьшается, но уменьшается их сила. (Рис.4).
Рис.4. Действие раздражения вагуса на кишечник (а, б, в) и сердце (г).
У беспозвоночных возбуждение в неисчерченных соматических мышцах (как с длинными, так и с короткими волокнами) проводится, вероятно, всегда по нервным волокнам. Широко распространена множественная моторная иннервация мышц, как исчерченных, так и неисчерченных. У позвоночных некоторые неисчерченные мышцы (например, мигательная перепонка) требуют нервного проведения; в других (гладкие мышцы внутренних органов) возбуждение проводится непосредственно от волокна к волокну. О висцеральных мышцах беспозвоночных известно очень мало, но в одной из них (веретенообразная мышца Phascolosoma) возбуждение от волокна к волокну проводится посредством механического растяжения.
Передача возбуждения в нервно-мышечных синапсах осуществляется химическими медиаторами, которые, за исключением ацетилхолина, норадреналина и 5-окситриптомина, еще не идентифицированы. Мембрана нервно-мышечного синапса обычно отличается по своим электрическим свойствам от проводящей мембраны даже той же самой мышечной клетки.
Раздражители, которые воздействуют на механорецепторы, рецепторы равновесия и фонорецепторы, занимают на шкале частот механических колебаний диапазон, простирающийся от постоянного давления через область кратковременных деформаций или низкочастотных вибраций до области ультразвуковых колебаний. Механорецепторы подразделяются на тактильные и проприоцептивные.
Говоря о механическом раздражении, не следует ограничиваться рассмотрением специфических рецепторов, поскольку большинство (а может и все) возбудимых клеток чувствительно к деформации поверхности. Существуют два общих типа механорецепторов: во-первых, те, у которых при механической деформации раздражается часть сенсорного нейрона, и, во-вторых, те, у которых при раздражении в эпителиальных клетках возникают электрические токи, в свою очередь возбуждающие нервные волокна, связанные с этими клетками. В обоих типах рецепторов возбуждение сенсорных аксонов имеет, по-видимому, электрическую природу.
Некоторые механорецепторы проявляют «спонтанную» активность. Следовательно, в центральную нервную систему непрерывно поступают сигналы. В таких органах чувств реакция на раздражитель выражается в изменении прежнего уровня активности, т.е. либо в повышении его, либо в понижении. Это изменение хорошо видно на рецепторах органов равновесия у рыб. Другие механорецепторы не проявляют активности в состоянии покоя. К таким рецепторам относится большинство тактильных волосков, а также имеющие капсулы рецепторы, реагирующие на прикосновение. Механорецепторы независимо от наличия или отсутствия у них спонтанной активности часто находятся под влиянием рефлекторной регуляции со стороны центральной нервной системы.
Мембраны механорецепторов сильно отличаются друг от друга по характеру возбуждения: одни из них в ответ на данное раздражение дают однократный разряд, другие – многократный.
Эхолокацию предметов на расстоянии, по-видимому, применяют некоторые водные насекомые, в небольшой степени рыбы (с помощью органов боковой линии), вероятно, китообразные.
Важным механизмом регуляции активности в нервных центрах служит реципрокное возбуждение и торможение. Один и тот же афферентный путь может быть возбуждающим для одной группы мотонейронов и тормозным для мотонейронов антагонистического действия. Тормозные нейроны, очевидно, действуют, высвобождая какой-то химический передатчик, и один и тот же передатчик может быть тормозным для одного нейрона и возбуждающим для другого.
По-видимому, можно считать всеобщим правилом, что торможение заключается в гиперполяризации, т.е. понижении мембранного потенциала или подавлении возбуждающей деполяризации. В сложных путях рефлекторных реакций происходит отставленное торможение с гораздо большим латентным периодом. Другим механизмом отставленного торможения может служить нейронная цепь с таким распределением процессов во времени, что импульсы, приходящие по определенному пути в соответствующее время, создают рефрактерность для импульсов, приходящих по другим путям.
Работа над ошибками по замечаниям на вопрос №3
Возбудимость – способность живых клеток, органов и целостных организмов воспринимать воздействия раздражителей и отвечать на них реакцией возбуждения. Возбудимость связана со специфической чувствительностью клеточных мембран, с их свойством отвечать на действие адекватных раздражителей (например: химических, механических) специфическими изменениями полной проницаемости и мембранного потенциала.
Возбуждение – реакция живой ткани на раздражения, характеризующаяся совокупностью физического, физико-химического и функционального изменения в ней. Во время возбуждения живая система переходит из состояния относительного покоя и деятельности, свойственной данной клетке или ткани (мышечная ткань – сокращается, нервная ткань – проводит возбуждение, железистая ткань – выделяет секрет).
В нервных и мышечных тканях возбуждение сопровождается появлением потенциала действия, способного без затухания распространятся вдоль всей клеточной мембраны.
Признаки возбуждения:
1. Потенциал действия – возникает ток действия;
2. Изменяется обмен веществ;
3. Изменяется температура.
Порог раздражения, порог возбудимости, минимальная интенсивность раздражения, способная вызвать распространяющийся потенциал действия, мере возбудимости клетки или организма в целом. Порог раздражения зависит от силы и качества раздражителя, длительности его воздействия и градиента нарастания силы, а также от свойств и физического состояния возбудимой ткани в момент раздражения. Сила раздражения ниже пороговой, т.е. не вызывающая раздражения называется подпороговой, а больше пороговой – надпороговой.
Раздражимость – способность живых клеток, тканей или целого организма реагировать на внутреннее или внешнее воздействие – раздражители; лежит в основе их приспособления к изменяющимся условиям внешней среды. Раздражимость проявляется на всех уровнях развития жизни и сопровождается комплексом неспецифических изменений, выражающихся в сдвигах обмена веществ, электрического потенциала, состоянии протоплазмы, а у высокоорганизованных животных с выполнением специфических функций (провидение нервного импульса, сокращение мышцы, выделение секрета железистой тканью и т.д.)
Раздражитель – стимул, любое воздействие, способное вызвать биологическую реакцию живой ткани, изменениями ее структуры и функции. Реакция ткани на раздражители называется раздражением.
Внешние раздражители – разные изменения внешней среды - световые и звуковые волны, химические и механические раздражения, действующие на клетки и органы чувств.
Внутренние раздражители – изменения состава и физических свойств жидких сред организма, а также степени наполнения внутренних органов.
Торможение – активный нервный процесс, результатом которого является ослабление или подавление процесса возбуждения. Различают периферическое торможение, осуществляемое непосредственно в синапсах на мышечных и железистых клетках и центральное, реализуемое в пределах ЦНС.
Список используемой литературы
Ихтиология. Моисеев П.А.; Азизова Н.А.; Куранова И.И. / М. ЛиПП 1981. - 384с.
Ихтиология. Баклашова Т.А. / М. Пищевая промышленность 1980. – 324с.
Практикум по ихтиологии. Скорняков В.И. и др. / М. Агропромиздат 1986. - 270с.
Физиология рыб. Методические указания. Аминева В.А. д.м.н.; Кузмин С.Ю. к.б.н. Калнг. «КГТУ» 2003. – 30с.
Физиология рыб. Аминева В.А.; Яржомбек А.А. / М. Легкая и пищевая промышленность 1984. -200с.
Поведение рыб. Протасов В.Р./ М. Пищевая промышленность 1978. -296с.
Сравнительная физиология животных. Под ред. Проссера Л./ М. Мир 1977. Т.2 -571с.; 1978. Т.3 -653с.
Справочник по физиологии рыб. Яржомбек А.А.; Лиманский В.В.; Щербина Т.В.; Бекина Е.Н.; Лысенко П.В./ М. Агропромиздат 1986. -192с.
[9] Экологическая физиология рыб. Строганов Н.С./ М. Московский университет. 1962. Т.1 -444с.
[10] Биологический энциклопедический словарь. Гиляров М.С./ М. Советская энциклопедия. 1986.

Нравится материал? Поддержи автора!
Ещё документы из категории биология:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ