Изучение биосинтеза аминокислот штаммом Вrevibacterium methylicum при росте на средах, содержащих тяжелую воду и дейтеро-метанол

 ИЗУЧЕНИЕ БИОСИНТЕЗА АМИНОКИСЛОТ ШТАММОМ Вrevibacterium methylicum ПРИ РОСТЕ НА СРЕДАХ, СОДЕРЖАЩИХ ТЯЖЕЛУЮ ВОДУ И ДЕЙТЕРО-МЕТАНОЛ.
ИЗУЧЕНИЕ БИОСИНТЕЗА АМИНОКИСЛОТ ШТАММОМ Вrevibacterium methylicum ПРИ РОСТЕ НА СРЕДАХ, СОДЕРЖАЩИХ ТЯЖЕЛУЮ ВОДУ И ДЕЙТЕРО-МЕТАНОЛ.
О. В. МОСИН
Московская государственная академия тонкой химической технологии им. М.В. Ломоносова, г. Москва, просп. Вернадского, д.86.
Представлены данные по биосинтезу дейтерий-меченных аминокислот L-фенилаланин-продуцирующим штаммом факультативных метилотрофных бактерий B. methylicum. Аминокислоты разной степени изотопной замещенности на дейтерий, как секретируемые в культуральную жидкость в процессе ферментации штамма-продуцента, так и в составе гидролизатов суммарных белков биомассы, были получены за счет биоконверсии СН3ОН/CD3OD на средах, содержащих различные концентрации тяжелой воды, включая 24,5 об. % D2O вплоть до 98 об.% D2O. Для изучения степеней включения дейтерия в аминокислоты использовали метод масс-спектрометрии электронного удара метиловых эфиров N-диметиламино-5-нафталин-сульфонильных (дансильных) производных аминокислот. Степени изотопного включения дейтерия в фенилаланин, секретируемый в культуральную жидкость и в составе гидролизатов суммарного белка изменяются от 27,5 до 95 % в зависимости от концентрации D2O в среде. Показано, что при ферментации наряду с основным продуктом биосинтеза (фенилаланин) в культуральной жидкости накапливаются другие аминокислоты , такие как аланин, валин и лейцин (изолейцин).
ВВЕДЕНИЕ
Метод мечения стабильными изотопами является ключевым направлением в разнопрофильных биохимических исследованиях с использованием аминокислот и других биологически активных соединений (БАС) [1,2]. Тенденции к предпочтительному применению стабильных изотопов, в частности, дейтерия по-сравнению с радиоактивными аналогами обусловлены такими их преимуществами, как отсутствием радиационной опасности и возможностью определения локализации метки в молекуле прямыми методами. Интенсивное развитие изотопно-чувствительной техники, прежде всего спектроскопии ядерного магнитного резонанса (ЯМР), инфракрасной и лазерной спектроскопии и масс-спектрометрии (МС), за последние годы позволило значительно усовершенствовать проведение многочисленных биологических исследований de novo, а также изучать структуру и механизм действия клеточных БАС на молекулярном уровне [3,4].
Начиная с первых экспериментов Катца с соавт. [5] по культивированию микроводорослей Chlorella на тяжелой воде продолжаются разработки методических подходов по получению БАС, в том числе аминокислот, меченных дейтерием. Потребности в дейтерированных аминокислотах обусловлены их важной ролью в разнообразных биохимических и медицинских исследованиях, а также возможностью их использования в синтезах широкого круга дейтерированных БАС, например, в синтезах пептидных гормонов и нейротрансмиттеров с использованием L-фенилаланина [6-8].
В настоящее время биотехнологический потенциал метилотрофных бактерий для получения аминокислот, меченных стабильными изотопами, в частности, дейтерием, общепризнан [9]. Современные методы мутагенеза и селекции открывают широкие перспективы для получения новых С1 -утилизирующих штаммов-продуцентов аминокислот, пригодных для роста на D2O. В связи с этим, большой практический интерес представляет исследования процессов биосинтеза аминокислот генетически маркированными штаммами метилотрофных бактерий, которые устойчивы к высоким концентрациям дейтерия в ростовых средах.
Раннее нами была изучена возможность использования штамма факультативных метилотрофных бактерий B. methylicum для получения дейтерированного фенилаланина [10]. В отличие от традиционных штаммов-продуцентов фенилаланина, у которых нарушены активности префенатдегидратазы или дезоксиарабиногептулозофосфатсинтетазы, уникальность этого штамма состоит в том, что для биосинтеза L-фенилаланина необходим L-лейцин.
Целью настоящей работы было изучение уровней включения дейтерия в молекулы аминокислот штамма B. methylicum как секретируемых в культуральную жидкость, так и аминокислот в составе белковых гидролизатов биомассы этих бактерий на средах с различным содержанием СН3ОН/CD3OD и D2O в них.
УСЛОВИЯ ЭКСПЕРИМЕНТА.
Бактериальные штаммы. Исследования проводили с L-лейцин-зависимым штаммом факультативных метилотрофных бактерий B. methylicum, продуцентом L-фенилаланина. Штамм был получен из коллекции культур Всероссийской коллекции промышленных микроорганизмов (ВКПМ) Государственного научно-исследовательского института генетики и селекции промышленных микроорганизмов.
Для приготовления питательных сред с различным содержанием дейтерия в них (см. табл.1) использовали безводные соли квалификации “х.ч.”, D2O (99,9% D) и СD3ОD (97,5 %D), полученные из Российского научно-исследовательского центра “Изотоп” (Санкт-Петербург, РФ). По необходимости тяжелую воду очищали от вредных примесей, перегоняя ее над перманганатом калия. Для получения производных аминокислот использовали N-диметиламинонафталин-5-сульфохлорид (дансилхлорид) (Sigma, CША) и диазометан. Диазометан получали из N-нитрозометилмочевины (Мerck, Германия).
Культивирование штамма В. methylicum проводили на минимальных средах М9 [11] как описано в работе [10].
Экстракцию липидов проводили смесью хлороформ-метанол (2:1) по методу Блайя и Дайера [12].
Щелочной гидролиз белка. Белок (4-5 мг) гидролизовали в запаянных стеклянных ампулах в 5 мл 4 н. Ba(OH)2 в течение 24 ч при 110 0С. Гидролизаты суспендировали в 20 мл горячей дист. воды и нейтрализовали 2 н. раствором H2SO4 с последующим отделением осадка центрифугированием (10 000 об/мин, 5 мин). Гидролизаты упаривали в роторном испарителе при 400 С.
Получение дансиламинокислот культуральной жидкости. К 200 мг лиофилизованных препаратов культуральной жидкости в 5 мл 2 м. NaHCO3 (2 10-3 моль) рН 9-10 дробными порциями при перемешивании добавляли 320 мг (1,2 10-3 моль) дансилхлорида в 5 мл ацетона. Реакционную смесь выдерживали при перемешивании при 400 С в течении часа, затем подкисляли 2 м. раствором HCL до рН 3,0 и экстрагировали этилацетатом (3 раза по 5 мл). Объединенный экстракт промывали водой до значения рН 7,0, сушили безводным сульфатом натрия, растворитель удаляли при 10 мм. рт. ст.
Дансиламинокислоты в составе белковых гидролизатов B. methylicum получали как описано в работе [9].
Получение метиловых эфиров дансиламинокислот. К 20 мл 40 %-ного КОН в 40 мл эфира добавляли 3 г влажной нитрозометилмочевины и перемешивали на водяной бане со льдом в течении 15-20 мин. После интенсивного газовыделения эфирный слой отделяли и промывали ледяной водой до рН 7,0, сушили безводным NaSO4 и обрабатывали им препараты дансилпроизводных аминокислот в составе культуральной жидкости и гидролизатов белка биомассы.
Количественное определение L-фенилаланина в культуральной жидкости проводили на спектрофотометре“Beckman DU-6” (США) при 540 нм после обработки препаратов культуральной жидкости нингидрином, как указано в работе [10].
Масс-спектры электронного удара производных аминокислот получены на приборе “MB-80A” (Hitachi, Япония) при энергии ионизирующих электронов 70 эВ.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ.
Получение дейтерий-меченных аминокислот и их масс-спектрометрический анализ.
Данные по росту штамма B. methylicum и максимальному уровню накопления фенилаланина в культуральной жидкости на средах с 2 об.% СН3ОН (СD3OD), содержащих ступенчато увеличивающиеся концентрации D2O представлены в таблице 1. Как видно из данных таблицы, рост исследуемых бактерий на средах с возрастающими концентрациями тяжелой воды сопровождался изменением выходов биомассы, времени клеточной генерации и продолжительности лаг-фазы при сохранении способности синтезировать и накапливать фенилаланин в культуральной жидкости. Поэтому было необходимо изучить, как изменяются степени включения дейтерия в фенилаланин и аминокислотные остатки суммарных белков биомассы B. methylicum в этих условиях.
ТАБЛИЦА 1.
Влияние изотопного состава среды на рост штамма B. methylicum и уровень накопления L-фенилаланина в культуральной жидкости.
Номер Компоненты среды, об% Величина Выход Время ген.
опыта лаг-фазы биомассы часы секреции
Н2О 2Н2О СН3ОН С2Н3О2Н часы % L-Phe,
1
98
0
2
0
24,0
100
2,2
100
2
98
0
0
2
30,3
92,3
2,4
99,1
3
73,5
24,5
2
0
32,1
90,6
2,4
96,3
4
73,5
24,5
0
2
34,7
85,9
2,6
97,1
5
49,0
49,0
2
0
40,5
70,1
3,0
98,0
6
49,0
49,0
0
2
44,2
60,5
3,2
98,8
7
24,5
73,5
2
0
45,8
56,4
3,5
90,4
8
24,5
73,5
0
2
49,0
47,2
3,8
87,6
9
0
98,0
2
0
60,5
32,9
4,4
79,5
10
0
98,0
0
2
64,4
30,1
4,9
71,5
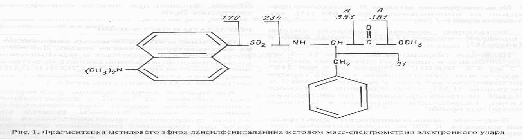
Для определения степени включения дейтерия в аминокислоты методом масс-спектрометрии электронного удара использовали метиловые эфиры дансил-аминокислот, которые химически стабильны и дают интенсивные пики молекулярных ионов в масс спектрах [13]. В качестве примера на рис.1 показана фрагментация метилового эфира дансилфенилаланина при электронном ударе. В масс-спектрах этого производного, как правило, четко детектируется пик молекулярного иона метилового эфира дансилфенилаланина М+. с m/z 412. Пик аминного фрагмента А имеет невысокую интенсивность, а пик аминоацильного фрагмента В крайне низкую или вообще отсутствует (см. рис. 1). Интенсивности пиков аминных А и аминоацильных В фрагментов других производных аминокислот, например присутствующих в культуральной жидкости аланина, валина и лейцина (изолейцина) при электронном ударе также сопоставимы. Кроме вышеобозначенных пиков, в масс-спектрах электронного удара метиловых эфиров дансил-аминокислот фиксируются пики с m/z 250, 234, 170, которые соответствуют дансильному фрагменту и продуктам его распада до N-диметиламинонафталина.
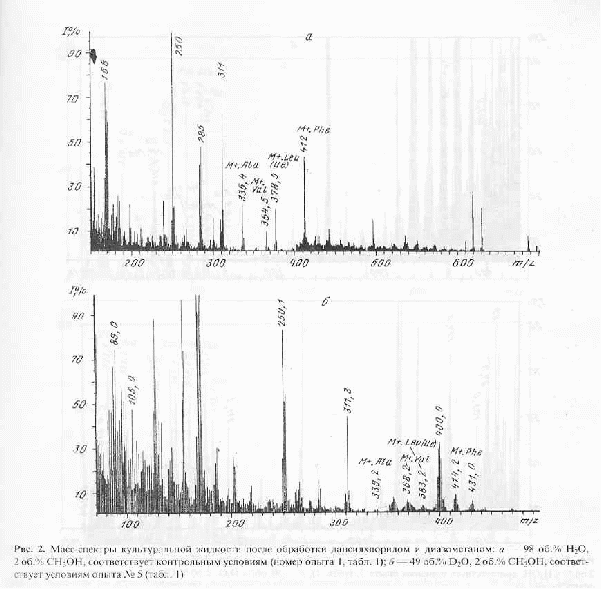
Модификация метода получения метиловых эфиров дансил-аминокислот заключалась в прямой обработке препаратов культуральной жидкости, полученной после отделения клеток, дансилхлоридом и диазометаном. В результате применения этого подхода удалось получить чётко интерпретируемые масс-спектры электронного удара метиловых эфиров дансил-аминокислот в присутствии следовых количеств низкомолекулярных метаболитов среды и сопутствующих аминокислот не прибегая к их предварительному хроматографического разделению и очистке. Следует отметить, что в условиях проведения дансильной дериватизации, для основной аминокислоты лизина характерно образование ди-дансильных производных, а для тирозина также происходит дополнительное дансилирование по гидроксильной -ОН-группе. При этерификации аминокислот диазометаном не исключено дополнительное N-метилирование по а-NH2-группе аминокислот, что приводит к появлению в масс-спектре метиловых эфиров дансил-аминокислот пиков, соответствующих соединениям с молекулярной массой на 14 массовых единиц больше исходных [14]. В качестве примера на рис. 2,б приведен масс-спектр секретируемого фенилаланина и сопутствующих аминокислот в составе дериватизированной культуральной жидкости в условиях эксперимента 5 (см. табл.1, опыт 5), где концентрация D2О в среде достигает 49 об.% (спектр приведен относительно контрольных условий (а), где использовали обычную воду и метанол). Из рис.2,б видно, что в этих условиях величина пика молекулярного иона производного фенилаланина (М+. с m/z 414,2) увеличивается по сравнению с контрольными условиями (М+. с m/z 412,0) на 2,2 единицы, что составляет 27,5 % от общего количества атомов водорода в молекуле. Пик с m/z 400, зафиксированный в масс-спектре культуральной жидкости (рис.2,б) вероятнее всего соответствует продукту отщепления метильной группы -СН3 от дейтерированного производного фенилаланина.
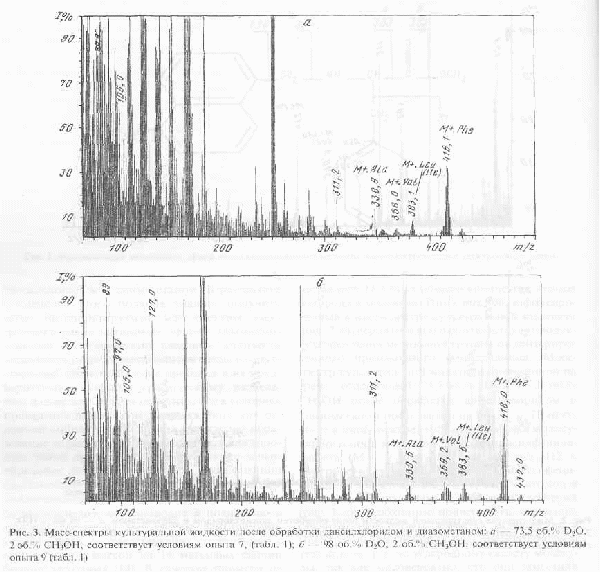
Масс-спектр культуральной жидкости, полученной на среде, содержащей 73,5 об.% D2О и 2 об. % СН3ОН после обработки дансилхлоридом и диазометаном представлен на рис. 3,а. Присутствие в масс-спектре этого образца пика молекулярного иона метилового эфира дансил-фенилаланина с М+. с m/z 416,1 (вместо 412 в контроле) указывает на увеличение массы фенилаланина на 4,1 единицу, т.е., 51,2 % атомов в молекуле фенилаланина замещены на дейтерий (рис. 3,а). Необходимо подчеркнуть, что вышеобозначенные атомы дейтерия включаются в молекулу фенилаланина за счет процесса биосинтеза de novo, т. е. по углеродному скелету молекулы, так как маловероятно, что они заместились в в ходе выделения аминокислоты из культуральной жидкости или при химической модификации фенилаланина. Что касается протонов (дейтеронов) при гетероатомах в NH2-, NH-, и -COOH группах аминокислот, то они за счёт лёгкости диссоциации легко обмениваются на дейтерий даже при растворении аминокислот в D2O. Так как все этапы, связанные с обработкой культуральной жидкости, содержащей дейтерий и её химической модификации проводились с использованием немеченных компонентов, растворителей и в т.ч. водных растворов, то эти протоны(дейтероны) из-за лёгкости диссоциации труднее всего поддаются точному подсчёту и контролю. Это могло служить причиной небольшого расхождения результатов при подсчёте степеней дейтерированности аминокислот.
Во всех исследуемых образцах культуральной жидкости B. methylicum кроме основной секретируемой аминокислоты (фенилаланин), обнаружены примеси (на уровне 3-5 мМ) метаболически связанных с ним аланина, валина и лейцина (изолейцина) (см., например рис 3,а). Как видно из рис.3,а, изотопный состав аланина характеризовался увеличением молекулярной массы на 2,5 единицы, валина-3,5 единицы, а лейцина (изолейцина)-4,6 единицами. Таким образом, в отличие от фенилаланина, количество включенного дейтерия в последних трех аминокислотах сохраняет стабильное постоянство в довольно широком интервале концентраций D2O в среде (от 49 об.% до 98 об.%).
Контроль за включением дейтерия в фенилаланин за счет ассимиляции дейтерометанола при росте бактерий на среде, содержащей обычную воду и 2% СD3OD (соответствуют опыту 2, табл.1) показал незначительное количество дейтерия, которое поступает в молекулу фенилаланина вместе с углеродом СD3OD. Процент дейтерирования фенилаланина был оценен по величине пика 413 за вычетом вклада пика примеси природного изотопа (не более 4 %, масс-спектр не приведён). Полученный результат может быть обусловлен разбавлением дейтериевой метки за счёт протекания как биохимических процессов, связанных с распадом дейтеро-метанола при его усвоении клеткой, так и реакциями изотопного обмена и диссоциации. Например, из четырёх атомов дейтерия, имеющихся в молекуле СD3OD, лишь атом дейтерия при гидроксильной группе --OD самый подвижный и поэтому легко диссоциирует в водной среде с образованием СD3OH. Три оставшихся атомов дейтерия в составе СD3OH входят в цикл ферментативного окисления метанола, который также мог привести к потере дейтерия за счёт образования соединений более окисленных, чем метанол. В частности, полученный результат по уровню включения дейтерия в фенилаланин подтверждает классическую схему ферментативного окисления метанола до формальдегида в клетках метилотрофов, который лишь после этого ассимилируется у данного штамма метилотрофных бактерий рибулозомонофосфатным путем фиксации углерода [15].
Так как базовый штамм продуцент фенилаланина был ауксотрофом по лейцину, то очевидно, что уровни включения дейтерия в секретируемый L-фенилаланин на фоне максимальных концентраций тяжелой воды, могут быть ниже теоретически допустимых, вследствие функционирования в клетке ряда биохимических реакций, связанных с ассимиляцией протонированного L-лейцина извне. Отмеченная особенность проявляется при биосинтезе L-фенилаланина на дейтерированной среде, в которой помимо метанола источником дополнительных протонов является немеченный лейцин. Так, в фенилаланине, полученном со среды, содержащей 98 об.% D2О и 2 об.% СН3ОН, только шесть атомов (из восьми обсуждаемых) в молекуле биосинтетически замещены на дейтерий (М+. с m/z 418 вместо 412) (Масс-спектр приведён на рис. 3,б). Эти два незамещенные атома водорода могли происходить из лейцина и метанола, однако авторы не исключают, что подобный эффект является результатом не ассимиляции протонированных субстратов, а вкладом протонов солей в составе ростовой среды. Следует подчеркнуть, что в этом масс-спектре фиксируется пик обогащённого дейтерием бензильного фрагмента с m/z 97 (вместо 91 в контроле), что указывает на то, что местами локализации атомов дейтерия в молекуле фенилаланина являются положения С1-С6 ароматических атомов и сопредельное с ними положение при углеродном атоме b. Причем, как миниум четыре из них могут быть локализованы в самом бензольном кольце молекулы фенилаланина.
Масс-спектрометрический анализ смесей дейтерий меченных аминокислот белковых гидролизатов B. methylicum.
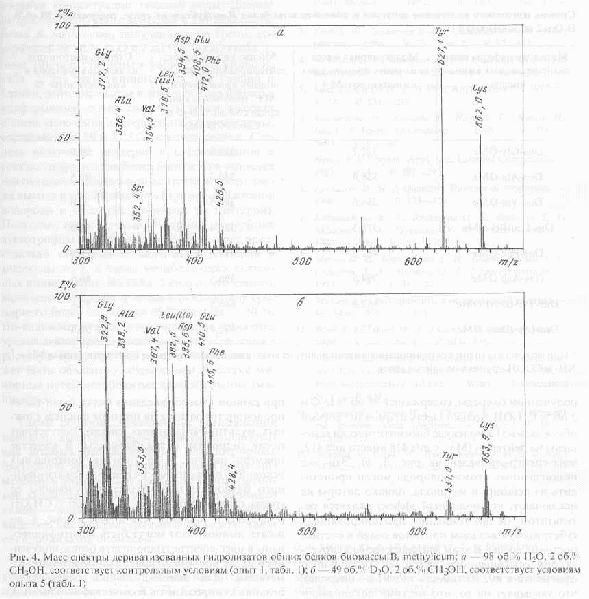
Общие принципы изучения степени дейтерированности аминокислот при данном способе введения метки были также продемонстрированы на примере анализа сложных мультикомпонентных смесей, полученных после гидролиза белка биомассы. В качестве примера на рис.4,б (относительно контрольных условий (а) приведен масс-спектр дериватизованного белкового гидролизата, полученного со среды, содержащей 49 об.% D2O и 2 об.% СН3ОН (табл.1, опыт (5)). Как видно из рис.4, до десяти аминокислот могут быть идентифицированы в масс-спектре гидролизата белка, полученного после обработки дансилхлоридом и диазометаном. Для индивидуальных аминокислот белковых гидролизатов количество включенных атомов дейтерия по скелету молекулы варьирует в пределах 49 %-ной концентрации D2O и составляет от 36,2 % для L-валина до 45,0 % для L-фенилаланина (рис. 4,б).
Вследствие того, что большой интерес представляет использование данного штамма метилотрофных бактерий для наработки униформно меченного белка и аминокислот, было необходимо изучить уровни включения дейтерия в аминокислоты белковых гидролизатов B. methylicum, при росте бактерий на среде, содержащей максимальные концентрации тяжелой воды. Данные по степеням включения дейтерия в аминокислоты белка B. methylicum, полученного со среды, содержащей 98 об.% D2O и 2 об.% СD3ОD (табл.1, опыт (10)) представлены в таблице 2. Как видно из таблицы 2, для таких аминокислот, как глицин и аланин дейтерирование в этих условиях близко к униформному, о чем свидетельствуют высокие степени включения дейтерия в эти аминокислоты, которые составляют 90 % и 97,5 % соответственно. Степень включения дейтерия в L-фенилаланин в составе гидролизатов белка биомассы в условиях максимально насыщенной дейтерием среды также высока, что составляет 95 % (примерно 8 атомов водорода в молекуле замещены на дейтерий). Низкие степени включения дейтерия в другие аминокислоты белка, прежде всего в лейцин (изолейцин) (49 %) в этих условиях могут быть объяснены за счет ауксотрофности штамма в лейцине, который добавляли в среду культивирования в протонированном виде. По-видимому, в условиях ауксотрофности по лейцину вклад атомов дейтерия, синтезируемых de novo в степень дейтерированности самого лейцина, а также метаболически близких с ним аминокислот незначителен.
ТАБЛИЦА 2.
Степени изотопного включения дейтерия в аминокислоты белка B. methylicum, полученного на среде, содержащей 98 об.% D2O и 2 об.% СD3OD.
Метиловые эфиры дансилпроизводных аминокислот
Молекулярные массы немеченных производных аминокислот, (М+.)
Молекулярные массы дейтерированных производных аминокислот (М+.), полученных на среде с 98 об% D2O и 2 об% CD3OD.
Степени изотопного включения дейтерия в аминокислоты, %
Dns-Gly-OMe
322,2
324,0
90,0
Dns-Ala-OMe
336,4
340,3
97,5
Dns-Val-OMe
364,5
368,5
50,0
Dns-Leu(Ile)-OMe
378,5
383,4
49,0
Dns-Phe-OMe
412,0
419,6
95,0
Dns-Asp-OMe
394,5
396,5
66,6
Dns-Tyr-(Dns)-OMe
662,0
668,5
92,8
Dns-Lys-(Dns)-OMe
627,1
632,4
58,9
Таким образом, проведенные исследования показали высокую эффективность использования штамма факультативных метилотрофных бактерий B. methylicum для получения дейтерий-меченных аминокислот разной степени изотопной замещенности на дейтерий, в том числе и униформно меченных. Эти аминокислоты можно выделять как из культуральной жидкости, так и из гидролизатов биомассы. Выбор штамма для этих исследований представляется авторам оправданным, так как B. methylicum характеризуется устойчивостью к максимальным концентрациям тяжёлой воды в среде.
ЛИТЕРАТУРА.
1. Beaufrere B, Fournier V, Salle B., Putet G. // American Journal of Physiology.- 1992 .- V.263. - N.1.- P.214-220.
2. Michalczuk L., Ribnicky D. M., Cooke T. J., Cohen J. D. // Plant Physiology. - 1992. - V.100. - N.3. - P.1346-1353.
3. Fesic S. W. and Zuiderweg E. R. // Quarterly Reviews of Biophysics. - 1990. - V.23. - N.2. - P. 97-131.
4. McIntosh L. P. and Dahlquist F. W. // Quarterly Reviews of Biophysics. - 1990. - V. 23. N.1. P. 1-38.
5. Katz J., and Crespi H. L. // Pure Appl. Chem. - 1972. - V.32. - P. 221-250.
6. Shimamura M., Kamada S., Hayashi T., Naruse H., Iida Y. // Journal of Chromatography. - 1986. - V.374. - N.1. - P. 17-26.
7. Hruby V. // J. Synth. and Appl. Isot. Labelled Compounds. - 1985. - V.4. - P. 287-292.
8. LeMaster D. M. // Quarterly Reviews of Biophysics. - 1990. - V.23. - P.133-174.
9. Karnaukhova E.N., Reshetova O.S., Semenov S.Y., Skladnev D.A., Tsygankov Y.D. // Amino Acids. - 1994. - V.6. - P.165-176.
10. Мосин О. В., Карнаухова Е. Н., Пшеничникова А. Б., Складнев Д. А., Акимова О. Л. // Биотехнология. - 1993. - Т.9. - С.16-20.
11. Миллер Дж. Эксперименты в молекулярной генетике. - М.: Мир, - 1976. - С. 393.
12. Bligh E.G., Dyer W.J. // Can. J. Biochem. Physiol. - 1959. - V.37. - N.8. - P.911-918.
13. Vetter W , in: Biochemical Applications of mass-spectrometry (Walles G.R., and Dormor O.C.). - 1980. - First supplementary volume. - Wiley - Interscience. - N.Y. - USA. - P.439.
14. Campbell J., Weiner W.C., Chess E.K. et al. // Biomedical inveron. mass spectrom. - 1990. - V.19. - P.520-522.
15. Nesvera J., Patek M., Hochmannova J. et al., Appl. Microbiol. 35, 777-780, (1991).
О.V. МОSIN
Moscow State Academy of Fine Chemical Technology named after М.V. Lomonosov, 117571.
STUDYING OF BIOSYNTHESIS OF AMINO ACIDS BY BACTERIAL STRAIN Вrevibacterium methylicum ON MEDIA, CONTAINING HEAVY WATER AND DEUTERO-METHANOL.
The data on biosynthesis of deuterated amino asids using the L-phenylalanine producing strain of facultative methylotrophic bacterium B. methylicum are presented. Amino acids with varying levels of isotope enrichment as secreted into culture medium during fermentation and amino acids derived from protein hydrolyzates of B. methylicum were obtained via bioconvertion of СН3ОН/CD3OD on media containing various concentrations of heavy water (including 24,5 v/v. D2O % up to 98 v/v. % D2O). For studying of levels of isotope enrichment the method of electrom impact mass-spectrometry of methyl esters of dansylamino acids was employd. The level of isotope enrichment of phenylalanine secreted into cultural medium as well as phenylalanine derived from protein hydrolizates was varried from 27,5% to 97,5 % (in dependence on biosynthesis conditions). It was shown, that during fermentation the amount of other amino acids such as alanine, valine, and leucine(isoleucine) can be produced and accumulated in culture medium in addition to the main product of biosynthesis (phenylalanine).

Нравится материал? Поддержи автора!
Ещё документы из категории биология:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ