Неспецифический иммунитет
 Содержание
Содержание
Введение
Актуальность
Врожденный неспецифический (естественный) иммунитет. Клеточный иммунитет
Фагоциты
Бактерицидная активность фагоцитов
Заключение
Список использованной литературы
Введение
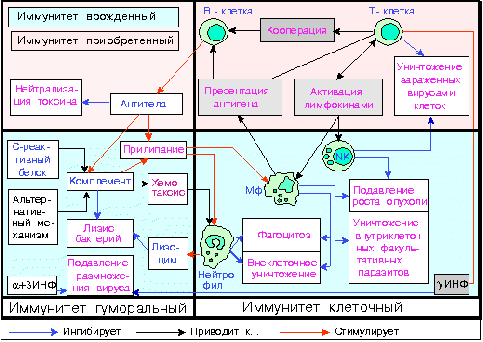
На протяжении многих миллионов лет существования жизни на Земле природа создала сложную, но надежную систему, получившую название иммунной или иммунокомпетентной. Основной задачей этой системы является создание условий для того, чтобы конкретный организм, конкретный индивидуум не погибал. Это значит, что под контролем иммунной системы находится функционирование очень многих органов и систем организма. Общую систему иммунитета можно разделить на два больших отдела, совместное функционирование которых создает очень мощную, имеющую несколько звеньев, защиту: врожденный неспецифический (естественный) иммунитет (неспецифические факторы естественной резистентности) и приобретенный специфический адаптивный иммунитет.
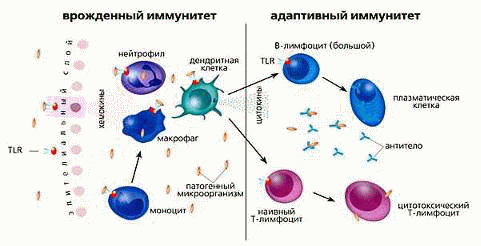
Если микроорганизму удается проникнуть через первичные барьеры, он сталкивается с клетками и механизмами системы врождённого иммунитета. Врождённая иммунная защита неспецифична, то есть её звенья распознают и реагируют на чужеродные тела независимо от их особенностей. Эта система не создает длительной невосприимчивости к конкретной инфекции. Система врождённого иммунитета осуществляет основную защиту у большинства живых многоклеточных организмов.
Актуальность
В настоящее время одной из наиболее актуальных проблем фундаментальной и клинической иммунологии является изучение врожденного иммунитета. Врожденный иммунитет является первой линией защиты организма от патогенов. Он реализуется через клеточные и гуморальные факторы. Факторы врожденного иммунного ответа предсуществуют или индуцируются быстро после инфекции. Компоненты врожденного иммунного ответа не изменяются в процессе жизни организма, контролируются генами зародышевой линии и передаются по наследству.
Система врожденного иммунитета - это самая первая линия обороны на пути инфекционного агента, пытающегося проникнуть в организм. Удивительные факты, обнаружившиеся при ее исследовании, помогут в создании новых методов лечения заболеваний и патологий, связанных с нарушениями иммунитета. [10]
Обязательным атрибутом нормальной аэробной жизни является генерация так называемых кислородных метаболитов [4]. Эти высокореактивные соединения обладают широким спектром биологического действия, в том числе проявляют свой мощный окислительный потенциал в бактерицидности фагоцитов.
Первоначально изучение активных кислородных радикалов, образующихся в ходе респираторного взрыва при фагоцитозе, было направлено на обнаружение механизмов возникновения кислородных радикалов, а также на выявление вклада продуктов неполного восстановления кислорода в бактерицидность фагоцитов по отношению к целому ряду микроорганизмов [9]. В последние годы наряду с супероксиданион-радикалом, перекисью водорода, гидроксильным радикалом, синглетным кислородом и гипогаллоидами, были обнаружены новые бактерицидные агенты фагоцитов - азотсодержащие метаболиты кислорода. В частности, показана важная роль оксида азота (NO), образующегося при взаимодействии азота аргинина с молекулярным кислородом, и пероксинитрита - продукта неферментативного взаимодействия NO - с супероксиданионом [8].
Кроме того, предметом пристального изучения являются механизмы и условия выживания бактериальной клетки при столкновении с кислородзависимыми бактерицидными факторами фагоцитов. При этом выживание бактерий при фагоцитозе наиболее часто связывается с выработкой бактериальными клетками ферментов-антиоксидантов (каталаз/пероксидаз и супероксиддисмутаз), которым нередко приписывается ведущая роль в обеспечении устойчивости к активным формам кислорода (АФК) [13].
Цели:
Обобщить представления о клеточных факторах защиты врожденного иммунитета.
Указать современные данные о кислородзависимых механизмах защиты макроорганизма, осуществляемых фагоцитирующими клетками.
Подробно описать пути образования реактивных метаболитов кислорода в клетках и ферментативные системы, принимающие участие в их наработке.
Охарактеризовать бактерицидные свойства реактивных метаболитов кислорода и их роль как физиологических медиаторов при воспалении.
Отметить различие в реализации бактерицидной активности нейтрофилов и макрофагов.
Проанализировать сведенья о роли нейтрофилов в кооперации фагоцитов при инфекциях и привести доказательства о способности этих клеток к синтезу и выделению низкомолекулярных БАВ.
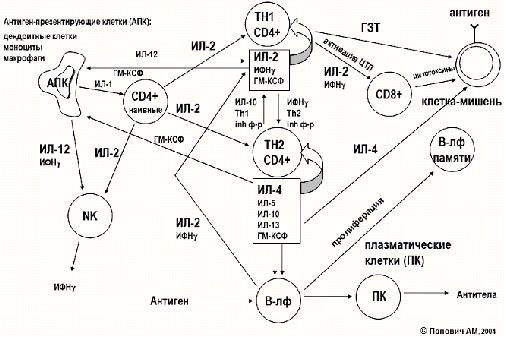
Врожденный неспецифический (естественный) иммунитет. Клеточный иммунитет
Это, прежде всего механические барьеры и физиологические факторы, которые препятствуют проникновению инфекционных агентов в организм: неповрежденная кожа, различные секреты, покрывающие эпителиальные клетки и предотвращающие контакт между разнообразными патогенами и организмом. К неспецифическим факторам резистентности можно отнести такие физиологические функции, как чихание, рвота, понос, которые также способствуют элиминации патогенных агентов из организма. Сюда же следует отнести такие физиологические факторы, как температура тела, концентрация кислорода, гормональный баланс. Этот последний фактор имеет большое значение для иммунного ответа. Далее можно выделить химические и биохимические реакции, подавляющие инфекцию в организме: продукты жизнедеятельности сальных желез, содержащие антимикробные факторы в виде жирных кислот, фермент лизоцим, который содержится в различных секретах организма и обладает способностью разрушать грамположительные бактерии; низкая кислотность некоторых физиологических секретов, препятствующих колонизации организма различными микроорганизмами.
Врожденный иммунитет можно разделить на клеточный и гуморальный.
К гуморальным факторам относятся:
лизоцим
система комплимента.
белки острой фазы.
цитокины и интерфероны.
белки теплового шока.
К клеточным факторам относятся:
фагоцитирующие клетки.
натуральные (естественные) клетки-киллеры (NK-клетки) [2].
вспомогательные клетки.
Вспомогательными клетками считаются тучные клетки, базофилы, эозинофилы, тромбоциты. Также в иммунной защите участвуют соматические клетки различных тканей организма. Тучные клетки находятся в соединительной ткани и слизистых оболочках и участвуют в регуляции воспалительной реакции. Они очень часто связаны с аллергией и анафилаксией. Они во многом напоминают базофилы - одну из малочисленных подгрупп зернистых лейкоцитов. Базофилы и эозинофилы родственны нейтрофилам. Эозинофилы секретируют биохимические медиаторы, которые участвуют в защите от крупных многоклеточных паразитов, а также играют роль в аллергических реакциях, например при бронхиальной астме.
NK (Натуральные (нормальные) киллеры (НК).
Общая характеристика Нормальные киллеры или NK (от англ. natural killer) - это большие зернистые лимфоциты с характерной морфологией: основная часть обильной цитоплазмы содержит несколько митохондрий, свободные рибосомы с отдельными элементами шероховатого эндоплазматического ретикулума, аппарат Гольджи и характерные электроноплотные гранулы, связанные с мембраной. Большие зернистые лимфоциты с активностью нормальных киллеров выполняют цитотоксические функции, также как и цитотоксические T - лимфоциты. Помимо механизма действия NK показан механизм действия цитотоксических T - лимфоцитов, которые прикрепляются к мишени в результате специфического узнавания поверхностного антигена, связанного с молекулами MHC класса I.
Основная обязанность киллерных клеток - выявлять и уничтожать собственные клетки организма, в которых что-то нарушилось: они убивают опухолевые клетки и клетки, зараженные вирусами (а также, возможно, и другими чужеродными агентами).
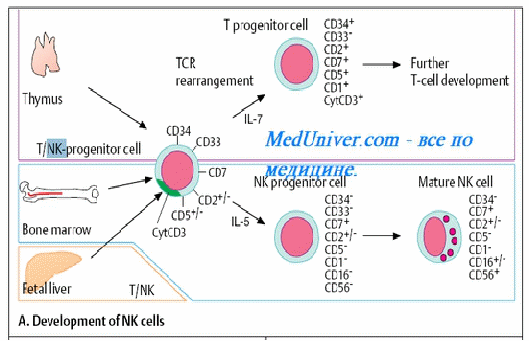
Нормальные киллеры (НК-клетки) у человека составляют примерно 5% лимфоцитов периферической крови. Чаще всего они имеют фенотип CD3-CD16+CD56+CD94+ и гаметное (неперестроенное) расположение генов. Таким образом, хотя НК относятся к лимфоидным клеткам, они лишены маркеров Т - и В-лимфоцитов.
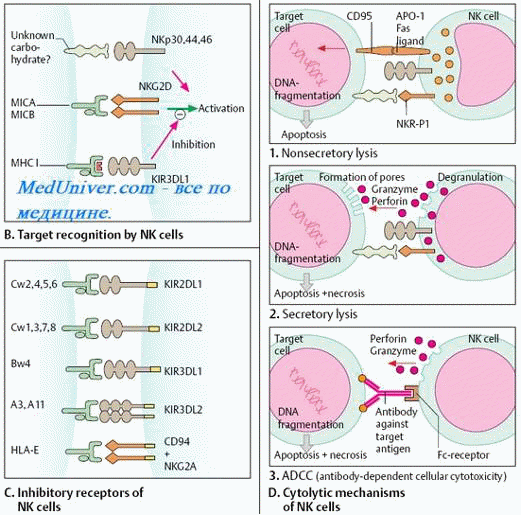
Полагают, что NK узнают определенные структуры высокомолекулярных гликопротеинов, которые экспрессируются на мембране инфицированных вирусом клеток. Узнавание клетки-мишени и сближение с ней происходит за счет рецепторов NK. В результате NK активируются, и содержимое гранул выбрасывается во внеклеточное пространство. Возможно, главная роль здесь принадлежит перфорину (цитолизину), имеющему некоторое структурное сходство с компонентом комплемента C9 (антитела к перфорину подавляют внеклеточное уничтожение). Перфорин встраивается в мембрану клетки - мишени и образовывает трансмембранные поры, что приводит к гибели клетки, поскольку содержимое клетки вытекает через эти поры. Кроме того, гранулы NK содержат две сериновые протеинкиназы, которые могут функционировать как цитотоксические факторы, но не вполне ясна их роль в NK-зависимом лизисе. Хондроитинсульфат A - протеогликан, устойчивый к протеинкиназам - тоже обнаружен в NK и может защищать эти клетки от автолиза [12].
NK-клетки не имеют основных маркёров Т - или B-лимфоцитов (поэтому их также называют нулевые лимфоциты), но экспрессируют дифференцировочные CD2, CD56 и CD16 (рецептор Fc-фрагмента AT) Аг. В отличие от цитотоксических лимфоцитов, способность NK-клеток к цитолизу связана с самостоятельным распознаванием "своё-чужое" на поверхности мишени [11].
При распознавании мишени НК-клетки способны как к "положительному", так и к "отрицательному" распознаванию.
В отличие от T-киллеров НК-клетки несут рецепторы подавления цитотоксичности ( KIR, англ. killer inhibitory receptor). При отрицательном распознавании, взаимодействуя с молекулами MHC класса I на клетке-мишени, эти рецепторы дают инфицированной клетке сигнал торможения ее цитотоксической активности. Положительное распознавание происходит, когда на клетках-мишенях отсутствует экспрессия молекул MHC, и взаимодействие НК-клеток с инфицированными клетками происходит с участием их собственных (НК-клеток) особых рецепторов, в частности CD2 и CD69, или антител, с которыми они связываются через рецептор для Fc ( CD16). Связывание НК с антителами, образовавшими иммунные комплексы с антигенами на поверхности клеток-мишеней, интерпретируется как проявление киллерной клеточной активности, или антителозависимой клеточной цитотоксичности. К примеру, вирусы герпеса пытаются избежать распознавание T-киллерами, подавляя экспрессию молекул MHC класса I на поверхности инфицированных клеток; однако в этом случае вирус распознают НК-клетки.
Следовательно, цитотоксические T-клетки (T-киллеры) и НК-клетки можно рассматривать как два взаимодополняющих инструмента иммунитета против вирусной инфекции тканей.
Набор клеток, подвергающихся литическому действию НК, достаточно широк. Это ряд вирусинфицированных и опухолевых клеток; клетки, на поверхности которых представлены цитофильные антитела; эмбриональные клетки.
NK-клетки уничтожают клетку-мишень после установления с ней прямого контакта при помощи специальных белков - перфоринов. Перфорины встраиваются в мембрану чужеродной или трансформированной клетки, образуя в ней "дыру", приводящую к необратимому и гибельному выравниванию ионного состава между цитоплазмой и внешней средой.
Активность NK-клеток регулируют цитокины (у-ИФН и ИЛ-2 усиливают их цитолитическую активность). Наряду с макрофагами, нейтрофилами и эозинофилами они также участвуют в антителозависимом клеточно-опосредованном цитолизе. Для этого NK-клетки экспрессируют на своей поверхности рецептор Fc-фрагмента IgG (GD16). Реакция зависит от присутствия AT, узнающих клетку-мишень и связывающихся с ней. Fc-фрагмент связанных с клеткой-мишенью AT взаимодействует с рецептором Fc-фрагмента, встроенным в плазматическую мембрану NK-клетки. Природа агента, убивающего клетку-мишень в этом случае, неизвестна [11].
Точное происхождение NK до сих пор остается неясным. Они происходят, в основном, из больших гранулярных лимфоцитов (БГЛ) . Несмотря на то, что НК морфологически напоминают лимфоциты или лимфобласты, их гистогенетическая связь с Т - или В-лимфоцитами не установлена. Вероятно, НК относятся к самостоятельной линии дифференцировки, хотя на самых ранних этапах развития у них имеется общий с лимфоцитами предшественник. В отличие от лимфоцитов, НК не имеют антигенраспознающих рецепторов, не увеличиваются количественно после взаимодействия с чужеродным (например, вирусным) антигеном и не способны к формированию иммунологической памяти. При этом их активность повышается под влиянием цитокинов Т-клеток и, в первую очередь, интерферона-гамма. Одной из характеристик НК является наличие Fc-рецептора. [12].
Вывод:
Нормальные киллеры или NK-клетки. Это лимфоциты, обладающие цитотоксической активностью, то есть способные прикрепляться к клеткам-мишеням, секретировать токсичные для них белки, убивать их или отправлять в апоптоз. NK-клетки распознают клетки, пораженные некоторыми вирусами, и опухолевые клетки. Они содержат на мембране рецепторы, реагирующие со специфическими углеводами поверхности клеток-мишеней [10].
Фагоциты
Фагоцитоз представляет собой важную особенность клеточного звена врождённого иммунитета, которую осуществляют клетки, называемые фагоцитами, которые "заглатывают" чужеродные микроорганизмы или частицы. Фагоциты обычно циркулируют по организму в поисках чужеродных материалов, но могут быть призваны в определённое место при помощи цитокинов [7] .
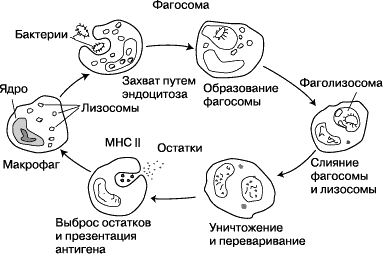
После поглощения чужеродного микроорганизма фагоцитом он оказывается в ловушке внутриклеточного пузырька, который называется фагосомой. Фагосома сливается с другим пузырьком - лизосомой, в результате чего формируется фаголизосома. Микроорганизм погибает под воздействием пищеварительных ферментов, либо в результате дыхательного взрыва, при котором в фаголизосому высвобождаются свободные радикалы. Фагоцитоз эволюционировал из способа получения захвата питательных веществ, но эта роль у фагоцитов была расширена, став защитным механизмом, направленным на разрушение патогенных возбудителей. Фагоцитоз, вероятно, представляет собой наиболее старую форму защиты макроорганизма, поскольку фагоциты обнаруживаются как у позвоночных, так и у беспозвоночных животных.
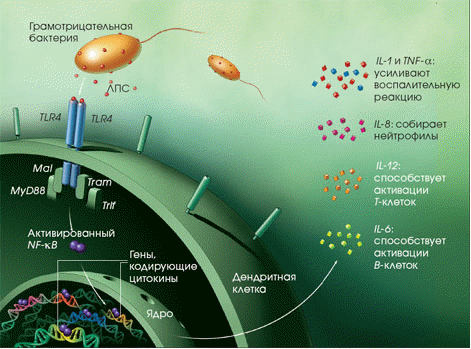
К фагоцитам относятся такие клетки, как мононуклеарные фагоциты (в частности - моноциты и макрофаги), дендритные клетки и нейтрофилы. Фагоциты способны связывать микроорганизмы и антигены на своей поверхности, а затем поглощать и уничтожать их. Эта функция основана на простых механизмах распознавания, позволяющих связывать самые разнообразные микробные продукты, и относится к проявлениям врождённого иммунитета. С появлением специфического иммунного ответа мононуклеарные фагоциты играют важную роль в его механизмах путём представления антигенов T-лимфоцитам. Для эффективного уничтожения микробов фагоцитам требуется активация.
Нейтрофилы и макрофаги представляют собой фагоциты, которые путешествуют по организму в поисках проникших сквозь первичные барьеры чужеродных микроорганизмов. Нейтрофилы обычно обнаруживаются в крови и представляют собой наиболее многочисленную группу фагоцитов, обычно представляющую около 50% -60% общего количества циркулирующих лейкоцитов. Во время острой фазы воспаления, в частности, в результате бактериальной инфекции, нейтрофилы мигрируют к очагу воспаления. Этот процесс называется хемотаксисом. Они обычно являются первыми клетками, реагирующими на очаг инфекции. Макрофаги представляют собой клетки многоцелевого назначения, обитающие в тканях и производящие широкий спектр биохимических факторов, включая ферменты, белки системы комплемента и регуляторные факторы, например интерлейкин-1. Кроме того, макрофаги выполняют роль уборщиков, избавляя организм от изношенных клеток и другого мусора, а также роль антиген-презентирующих клеток, активирующих звенья приобретённого иммунитета. [4]
Дендритные клетки представляют собой фагоциты в тканях, которые соприкасаются с внешней средой, то есть, расположены они, главным образом, в коже, носу, лёгких, желудке и кишечнике. Они названы так, поскольку напоминают дендриты нейронов наличием многочисленных отростков, однако дендритные клетки никоим образом не связаны с нервной системой. Дендритные клетки служат связующим звеном между врождённым и приобретённым иммунитетом, поскольку они представляют антиген T-клеткам, одному из ключевых типов клеток приобретённого иммунитета.
Бактерицидная активность фагоцитов
Еще в 1908 г. русский ученый И.И. Мечников открыл не только прямые бактерицидные свойства микрофагов-нейтрофилов и макрофагов, но и также предполагал другие возможные их функции, как, например, передача иммунитета…посредствам белых корпускул через продукцию "секретинов". Нейтрофилы являются короткоживущими, но чрезвычайно многочисленными, и им была отведена фундаментальная роль в разрушении внеклеточных патогенов и их токсинов, тогда как другая группа фагоцитов, производных от моноцитов-макрофаги, относятся к длительноживущим клеткам. В дополнение к бактерицидной активности фагоциты способны осуществлять ряд других важных функций, таких как: ограничение роста облигатных внутриклеточных патогенов, продукция многих биоактивных молекул, необходимых для регуляции различных функций клеток (компоненты комплемента, простагландины и цитокины), удаление дефектных клеток.
Первичный ответ на внедрение микробного патогена в организм осуществляется резидентными макрофагами, которые продуцируют факторы воспаления и изменяют хемотаксический градиент. В свою очередь, нейтрофилы отвечают на межклеточные сигналы и проходят через эпителий, при этом их слабая адгезия опосредована - селектинами. Эти фагоциты уже активизированы для адгезии к эндотелию посредством собственных мембранных интегринов и способны распознавать специальными рецепторами - серпентинами - специфические хемоаттрактанты. Одновременно с этим происходит изменение формы клетки посредством перегруппировки активного цитоскелета. Нейтрофилы продолжают получать информацию, которая в дальнейшем изменяет их ответную реакцию.
Известны два отчетливо различимых функциональных состояния фагоцитов "redox", с низким уровнем протекания процессов, и активированное, который обусловлен взаимодействием клеток с различными стимуляторами. При этом в процессе предварительного воздействия стимулов отмечается усиление миграции, адгезии, дегрануляции и метаболизма нейтрофилов и макрофагов. Это явление получило название прайминга, то есть подготовки, перевода клеток в рабочее состояние. Таким образом, нейтрофилы, достигнув места воспаления, способны распознать патоген либо непосредственно через мембранные рецепторы для опсонинов, либо через лектины микробов и фагоцитов. В дальнейшем начинается процесс фагоцитоза, осуществляемый с помощью механизма, действующего как замок молния. Происходит поглощение микробов посредством инвагинации плазматической мембраны клеток и образование фагоцитарной вакуоли. При этом параллельно активируется две функции фагоцитов: выброс содержимого гранул в фагосому и кислородный взрыв.
Феномен кислородного взрыва впервые был описан в 1933 г., его суть состоит в том, что фагоциты резко увеличивают потребление кислорода - от 50 до 100 раз. Этот процесс происходит при стимуляции комплекса НАДФ - оксидазы, также известной как фагоцитарная оксидаза, или оксидаза кислородного взрыва. Субклеточная локализация этого комплекса хорошо изучена в нейтрофилах. В макрофагах этот комплекс выявляется только на плазматической мембране. Ввиду того, что составляющие этого комплекса, не обнаружены на мембранах гранул макрофагов - эти клетки не способны продуцировать реактивные виды кислорода внутрифагосомально.
На первом этапе для образования из молекулы кислорода супероксидного аниона О2 - донором электронов является НАДФ - оксидазный комплекс. Этот комплекс включает 4 белковых компонента, молекулярные массы, которых вошли в их название - Р40 PHOX (PHOX расшифровывается как фагоцитарная оксидаза. В клетках состояния покоя - “redox” - три этих компонентов - P40 PHOX,P47 PHOX и P67 PHOX находятся в цитозоле в виде комплекса, а два других компонента - P22 PHOX и gp91 PHOX локализованы на плазматической мембране или на мембране секреторных везикул и азурофильных гранул, где они представлены в виде гетеродимерных флавогемопротеинов, известных как цитохром b558. При стимуляции клеток цитозольный компонент р67 PHOX фосфорилируется и выступает в качестве адаптера связи компонента р67 PHOX с цитохрома b558, а в свою очередь, фосфорилирование р40 PHOX вызывает конформационные изменения р67 PHOX для его полноценной связи с цитохромом [9]. Таким образом, собранный НАДФ - оксидазный комплекс способен передаче электронов от субстрата к кислороду посредством собственных электрон-несущих протеазных групп - флавинов, преобразуясь при этом в так называемый восстановленный НАДФ+ - комплекс. В процессе сборки НАДФ-комплекса участвуют также низкомолекулярные белки, связанные с нуклеотидом гуаназин 5’ - трифосфатом - Rac. В процессе активации фагоцита Rac1/2 связываются с гуаназин фосфатом (ГТФ) и мигрируют к плазматической мембране вместе с основным цитозольным комплексом. В это же время цитохром b558 и Rap 1А доставляются к клеточной поверхности посредством слияния мембран, таким образом, локализуя НАДФ - оксидазный комплекс внутри фагосомы. Это позволяет эффективно доставлять активные метаболиты кислорода для уничтожения поглощенной мишени. Отмечаются различные пути регуляции НАДФ - оксидазы нейтрофилов в зависимости от того, где она локализована на плазматической мембране или на мембране гранул. Активация НАДФ - оксидазы, локализованы на плазматической мембране регулируется сложным путем комплекса сигналов, опосредованных через мощный приток Са2+. В моноцитах/макрофагах в процессе фосфорилирования принимает участие фермент - протеиновая киназа.
Продукт восстановления НАДФ - оксидазного комплекса супероксидный радикал - О2 - является начальным материалом для продукции обширного ряда реактивных оксидантов, включая окисленные галогены, свободные радикалы и синглетный кислород. Эти окислители используются для уничтожения поглощенных микроорганизмов, но они также вызывают множественное разрушение окружающих тканей, поэтому их формирование должно быть отрегулировано.
Второй этап - преобразование супероксидного аниона О2 в следующий мощный окислительный компонент перекись водорода (Н2О2) - может происходить спонтанно или катализироваться супероксиддисмутазой. Спонтанная реакция происходит при рН=4.8, при которой появляется равное количество О2 и Н2О. При повышенном значении рН и преобладании О2 падает показатель спонтанной дисмутации этого радикала и включается процесс катализации супероксиддисмутазой. Перекись водорода сама по себе не является бактерицидной. Подобный эффект отмечается только при ее высокой концентрации. Таким образом, экзогенно образованный супероксид и перекись водорода не способны на прямую убивать бактерии. Более мощным по бактерицидному воздействию являются другие реактивные метаболиты кислорода, образованные из перекиси водорода. В фагоцитах существуют 4 потенциальных механизма преобразования перекиси водорода. Первый путь осуществляется с помощью реакции Фентона, при участии сульфата железа. Ее результатом является гидроксильный радикал - ОН. К преобразованию трехвалентного железа в двухвалентное может подключаться супероксидный анион. Эта реакция получила названия супероксид - управляемой реакции или реакции Хабер-Вейса.
Содержание свободных ионов железа в клетках довольно низкое, и связанны с белками - миелопероксидазой, которая также может выступать в качестве катализатора при образовании перекиси водорода. Формирование гидроксильного радикала из НОCl и супероксида изолированными нейтрофилами зависит именно от наличия этого фермента. В настоящее время путь образования ОН с этим ферментом носит название "миелопероксидазно-Н2О2-хлоридная система".
Гидроксильные радикалы оказывают повреждающее действие на бактерии. Они способны, действуя на SH-группы, гистидиновые и другие аминокислотные составляющие белков, вызывать их денатурацию, таким образом, дезактивируя ферменты. Помимо этого в нуклеиновых кислотах ОН разрушает углеводные мостики между нуклеотидами и разрывает цепи ДНК и РНК, что может являться причиной мутации и гибели бактерий. Однако, по ряду причин эти радикалы не на столько эффективны в бактерицидности, как может предполагать их высокая реактивность. К ним относят ограниченное действие этих соединений из-за пространства фагосом, а также они могут прореагировать с другими субстратами, не достигнув бактерий. Тем не менее, было обнаружено, что гидроксильные радикалы, произведенные системами, включающими хлориды, наиболее токсичны для бактерий.
Синглетный кислород также продуцируется нейтрофилами при взаимодействии гидроксильного радикала с НОСl. При использовании ловушки для синглетного кислорода в нейтрофилах обнаружено высокое преобразование кислорода в синглетную форму при участии миелопероксидазно-Н2О2-хлоридной системы.
Недавно был назван ещё один реактивный метаболит кислорода - продукт респираторного взрыва в нейтрофилах: озон (О3). Озон сам по себе является бактерицидным, но в комбинации с перекисью водорода он ещё более токсичен для микроорганизмов.
В настоящее время, активно исследуется ещё один метаболит - оксид азота, присутствие которого отмечается и в фагоцитах. Оксид азота (NO) является свободным радикалом (газовой молекулой), который продуцируется из молекулярного кислорода и гуанидинового нитрогена L-аргинина, собранного в L-цитруллине. Было показано, что NO включается в неспецифический иммунитет и частично в комплексный механизм тканевого повреждения как важный медиатор воспалительных процессов и апоптоза. При этом цитотоксическое/ цитопатическое действие усиливается благодаря способности NO вступать в реакцию с супероксидным радикалом, образуя пероксинитрит.
Необходимо также упомянуть, что митохондриальное кислородное фосфорилирование занимает главное место в продукции реактивных видов кислорода как побочных продуктов электронной транспортной цепи. Количество митохондрий в различных типах клеток неравнозначно, и в фагоцитах оно не большое. В нейтрофилах почти весь потребляемый кислород минует малочисленные митохондрии и идет на образование оксидантов для осуществления прямой бактерицидной активности этих клеток. В макрофагах же имеется большее количество митохондрий, система которых гипертрофируется при активации клетки. В зрелых макрофагах место фермента миелопероксидазы существует другая альтернативная система разрушения Н2О2 и других активных форм кислорода, состоящая из каталазы и глутатионпероксидазы. Особенность альвеолярных макрофагов, которая и определяет их функциональную предназначенность как ключевых клеток воспаления, в том, что в процессе созревания из моноцита в макрофаг в этих фагоцитах отмечается резкое снижение внутриклеточного количества азурофильных гранул, которые включают основной арсенал бактерицидных ферментов: миелопероксидазу, сериновые протеазы, катионные белки и лактоферрин, характерные для нейтрофилов. Часто в литературе при описании включений фагоцитов отождествляют понятия "лизосомы" и "азурофильные гранулы". Эти две клеточные органеллы по своему содержанию отличаются друг от друга. Для азурофильных гранул характерно наличие биологически активных веществ, обладающих четко выраженным бактерицидным эффектом, то для лизосом - преимущественно содержание ферментов, участвующих в процессе переваривания поглощенных объектов. Эти кислые гидролазы и фосфатазы; при этом, как известно, кислая фосфатаза является специфическим маркером лизосом.
В результате исследований последних лет было установлено, что оксид азота, супероксиды и другие реактивные метаболиты кислорода участвуют во множестве физиологических и патологических процессов как сигнальные медиаторы - проводники. Временное регулирование преобразования различных источников реактивных метаболитов кислорода проходит путем модификации функции каскада сигнальной трансдукции. Если при высокой концентрации свободные радикалы кислорода и их производные являются опасными для живых организмов и могут избирательно разрушать определенные клетки, то при умеренной концентрации эти соединения могут играть важную роль в качестве регуляторных медиаторов в процессах сигнализации различных физиологических функций. К ним относятся: регуляция сосудистого тонуса, осуществление мониторинга в контроле кислородной вентиляции, в продукции эритропоэза и др.
Доказано, что оксид азота и реактивные метаболиты кислорода могут вызывать апоптоз различных типов клеток.
В настоящее время схема взаимодействия фагоцитов с патогенами в значительной степени была модифицирована:
Отмечается более длительный период жизни нейтрофилов, особенно при воспалениях и инфекциях.
Выявлены новые рецепторы, способные к распознаванию иммуномодуляторов.
Способность этих клеток к синтезу различных БАВ: цитокинов, таких как фактор некроза опухоли, интерлейкины, колониестимулирующие факторы, комплементарные белки, дефенсины и др.
Нейтрофилы могут регулировать экспрессию генов постоянно и индукционно. Уточняется роль этих фагоцитов в специфическом иммунном ответе. В частности, нейтрофилы способны кооперироваться с профессиональными антигенпредставляющими клетками через предобработку и протеолиз антигенов. При этом они могут вызывать экспрессию молекул класса II на мононуклеарных клетках, а также представлять антиген для вирусоспецифических Т-лимфоцитов памяти. Эти свойства проявляются параллельно с увеличением активности миелопероксидазно - Н2О2-хлоридной системы, чьи производные действуют как иммуномодуляторы, обеспечивая дальнейшую связь между первичным и адаптивным иммунитетом. Другой регулирующий аспект нейтрофилов включает продукцию и экзоцитоз различных факторов, которые модулируют функции лимфоцитов, моноцитов и эозинофилов.
Херст в 1956г. впервые описал антимикробные белки - фагоцитины, которые являются необработанной белковой фракцией полиморфноядерных лейкоцитов. Это положило начало исследованиям биологически активных белковых компонентов, которые нейтрофилы способны секретировать в очаге воспаления.
Особый интерес при исследовании белков цитоплазматических гранул нейтрофилов (их более 40) вызывают неферментные бактерицидные белки с низкой молекулярной массой, которые обладают суммарным положительным зарядом и бактерицидным действием. Эти протеины способны играть роль медиатора воспаления, фактора проницаемости, стимулятора метаболических процессов и служить источником не специфических опсонинов при фагоцитозе, модулируя свертывание крови и стимулируя адгезию, хемотаксис и зависимый от комплемента лизис. Данные белки являются специфическим маркером нейтрофилов. К группе этих белков относится: протеины, повышающие бактерицидные возможности клеток, кателицидины, которые содержаться в неактивной форме в азурофильных гранулах нейтрофилов, протеазы, способные утилизировать большую часть компонентов межклеточного матрикса и другие протеины. Все они могут вступать в роли физиологических медиаторов.
Следующая важная группа антимикробных пептидов, которую нельзя не упомянуть, это дефенсины, низкомолекулярные катионные белки, содержащие 6 цистеиновых и дисульфидных связей. И их деятельность направлена против грамотрицательных и грамположительных бактерий и осуществляется посредством нарушения целостности микробных мембран. Помимо бактерицидности, дефенсины проявляют свойства регуляторов воспалительного процесса, связываясь с ингибиторами протеазы.
Вывод:
Таким образом, исходя из вышеизложенного, можно сказать, что нейтрофилы обладают богатым потенциалом низкомолекулярных БАВ, которые они способны выделять во внеклеточное пространство. Определенное место занимают и молекулярные межклеточные медиаторы, выделяемые нейтрофилами-активные метаболиты оксида азота и кислорода. Эти межклеточные посредники фагоцитов способны контролировать развитие воспаления, как на ранней, так и на поздней стадии иммунного ответа организма, и такая взаиморегуляция функциональной активности этих клеток может вытекать из факта присутствия в организме их общей родоначальной клетки [6].
Активность фагоцитов характеризуется фагоцитарными показателями и опсонофагоцитарным индексом. Фагоцитарные показатели оцениваются числом бактерий, поглощенных или "переваренных" одним фагоцитом в единицу времени, а опсонофагоцитарный индекс представляет отношение фагоцитарных показателей, полученных с иммунной, т.е. содержащей опсонины, и неиммунной сыворотки. Эти показатели используются в клинической практике для определения иммунного статуса индивидуума. [3].
Заключение
Таким образом, система врожденного иммунитета создает мощный заслон для инвазии бактерий, вирусов, грибков, простейших или многоклеточных паразитов и содержит эффективные механизмы для их распознавания, обезвреживания и удаления из макроорганизма [1].
В заключение, следует отметить, что и мононуклеарные фагоциты, и киллерные клетки относятся к первой линии защиты при попадании в организм инфекционного возбудителя [7].
Список использованной литературы
1. Абелев Г.И. Соровский Образовательный Журнал, 1998г.,№2.
2. Борисов Л.Б. медицинская микробиология, вирусология, иммунология. (2002), стр.217
3. Воробьева А.А. Медицинская микробиология, вирусология и иммунология (2006), стр. 197
4. Зенков Н.К., Меньщикова Е.Б. (1993)
5. Пиненгин Б.В., Дамбаева С.В. Журнал Иммунология №2, 2007г., лекция.
6. Плехова Н.Г. Журнал Микробиология, 2006г., №6, стр.89-96
7. http://immuno. health
8. Gryglewski R., Palmer R., Moncada S., 1986; Beckman J., Beckman Т., Chen J., 1990; Ischiropoulos H., Zhu L, Beckman J., 1992.
9. Allen R. С, Stjernholm R. L, Steele R. H., 1972; Babior В., 1978; Clark R., Klebanoff S., 1978; Samokyszyn V. W., Thomas C. E., Reif D. W., Saito M., Aust. D., 1988; Ginsburg I., Kohen R., 1995.
10. http://immunar.ru
11. meduniver.com/Medical/Microbiology.
12. http://www.biotech. spb.ru
13. [Fridovich I. 1989; Wendel A. 1988; McCord J. M., Fridovich I., 1969a, б; Klebanoff S. J., 1993].

Нравится материал? Поддержи автора!
Ещё документы из категории биология:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ