Отдел динофитовые водоросли

 Отдел динофитовые водоросли
Отдел динофитовые водоросли
В подавляющем большинстве представителей — это монадные формы, реже ризоподиальные, пальмеллоидные, коккоидные, нитчатые. Принадлежность неподвижных форм так же, как многих бесцветных сапротрофных и паразитных форм к динофитовым водорослям, легко устанавливается по совершенно уникальному строению клеточного ядра и по подвижным репродуктивным стадиям, повторяющим характерные признаки монадных представителей отдела. Обычно линзовидные хлоропласты чаще всего окрашены в бурый цвет благодаря преобладанию каротиноидов над хлорофиллами а тл с. Оболочка хлоропластов состоит из трех мембран, ламеллы, как правило, трехтилакоидные, опоясывающая ламелла отсутствует, генофор рассеянный. Пиреноиды различной формы: простой внутренний пиреноид, гранулированный матрикс которого не пересекается тилакоидами; сложный внутренний голый пиреноид, пронизанный более или менее параллельными ламеллами; пиреноиды, выступающие из хлоропластов в виде почки и сидящие на одной или многих ножках. Продукты ассимиляции — экстрапластидный крахмал и жир. Крахмал встречается преимущественно у пресноводных форм, жир — у морских.
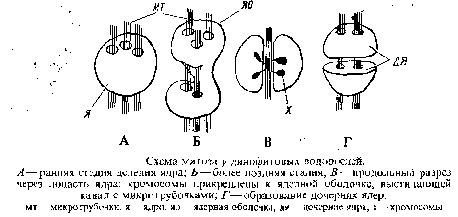
Ядерный аппарат. Все динофитовые водоросли имеют уникальное клеточное ядро, носящее специальное название динокарион, для которого характерно конденсированное состояние ДНК на протяжении всего митотического цикла, включая интерфазу. Таким образом, число хромосом может быть подсчитано на препаратах интерфазных ядер. Хромосомы имеют вид многочисленных палочковидных телец, имеющих исчерченную структуру, кинетохоры отсутствуют. Тесты с энзимами, такими, как ДНК-аза, в более ранних работах показали, что хромосомы динофитовых водорослей в отличие от хромосом эукариотических организмов состоят из ДНК-фибрилл без какого-либо заметного количества гистона. т. е. как у всех прокариотических организмов. Это побудило назвать динофитовые водоросли мезокариотическими организмами, занимающими по организации их ядра промежуточное положение между прокариотическими и зукариотическими водорослями. Позднее были представлены убедительные аргументы в пользу отнесения динофитовых к эукариотическим организмам, а недавно были обнаружены и низкие концентрации хромосомных белков. Таким образом, от термина мезо-кариотические следует отказаться. Ядра у различных представителей динофитовых водорослей помимо крупных размеров отличаются большим разнообразием формы: сферические, полукруглые, эллипсоидальные, четырехугольные, треугольные, почковидные. Они окружены ядерной оболочкой, как правило, с порами, и содержат ядрышко.
Митоз имеет ряд особенностей. Центриоли отсутствуют. При делении ядро увеличивается. Его оболочка остается интактной на протяжении всего митоза и возникают глубокие инвагинации ее внутрь ядра; некоторые из них пронизываю! ядро насквозь, образуя каналы или туннели. В этих цитоплазматических каналах, и таким образом вне ядра, возникают микротрубочки — эквивалент веретена. Сегрегация хроматид, возможно, осуществляется благодаря прикреплению их к ядерной оболочке, выстилающей туннели, которые заметно вытягиваются. В конце концов ядерная оболочка перетягивается по экватору и разделяет ядро и ядрышко, сохраняющееся на протяжении всего митоза, на две части.
Другие органеллы клетки — аппарат Гольджи, митохондрии — в общем, обычного строения. Аппарат Гольджи состоит из стопок уплощенных мембран, вздутых на концах, с многочисленными терминальными везикулами. Часто тела Гольджи располагаются по соседству с ядром. Митохондрии — вытянутые, иногда разветвленные
структуры, округлые на поперечном срезе, ограничены двойной мембраной; кристы короткие, трубчатые. Митохондрии рассеяны по цитоплазме. У Gonyaulax polyedra описано слияние отдельных митохондрий, приводящее к образованию гигантских митохондрий.
Монадные клетки динофитовых водорослей имеют ряд специфических особенностей, касающихся клеточного покрова, трихоцист, строения жгутикового аппарата, осморегуляторной системы клетки, фоторецепторного органа.
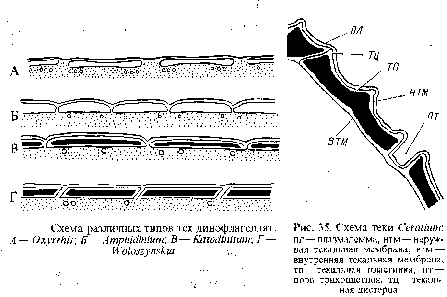
Клеточный покров. У монадных клеток динофитовых водорослей имеется сложный клеточный покров — тека или амфиезма, представленный цитоплазматической мембраной, под которой в один слой располагаются компоненты теки — уплощенные пузыри, окруженные одиночной мембраной. Число текаль-ных цистерн в клетке варьирует от нескольких сотен до двух. Расположение текальных везикул под плаз-малеммой неодинаково. Например, у рода оксирис они разъединены друг от друга, а у гимнодиниума, амфидиниума и других родов везикулы соприкасаются друг с другом у своих краев и приобретают многоугольные очертания. Везикулы у одних представителей, таких, как гимнодиниум, клетки которых в оптическом микроскопе выглядят как голые, не содержат текальпых пластинок, в них наблюдается только зернистое содержимое. У других в цистернах находятся целлюлозные пластинки, различающиеся по толщине. Имеется обратная зависимость между толщиной пластинок и их числом. Чем толще пластинки, тем их меньше. У катодиниума везикулы содержат тонкие пластинки, а у волошинскии пластинки достаточно толстые, чтобы их можно было различить в оптическом микроскопе. У родов гленодиниум, перидиниум, цератиум число пластинок редуцируется приблизительно до двадцати, они очень толстые и могут нести шипы, различные выросты, иметь сетчатый узор.
Пластинки вокруг клетки создают прочный целлюлозный панцирь. У пророцентрума в клетке только две главные толстые пластинки формируют целлюлозный панцирь. Во всех случаях в норме непосредственно под текой находятся микротрубочки.
В месте соприкосновения соседних текальных пузырей их мембраны располагаются очень тесно, образуя шов. Рост пластинок происходит в двух областях. Посредством отложения строительного материала у краев пластинок вдоль шва возникают ин-теркалярные полосы, обусловливающие плоскостной рост пластинок. У Gonyaulax polyedra вдоль мембран швов по обе стороны наблюдается электронно-плотный слой, который, возможно, принимает участие в плоскостном росте пластинок. В результате отложения материала вдоль внутренней и наружной сторон пластинок они утолщаются и на верхней стороне пластин развивается характерная для каждого вида скульптура. Необходимый для роста в толщину материал, возможно, доставляется в текальные пузыри с помощью специальных шаровидных везикул и от внутренней стороны пластин на их наружную сторону — через трихоцистные поры.
Иногда у отдельных клеток динофлагеллят наблюдается процесс сбрасывания панциря, который можно искусственно стимулировать и затем наблюдать регенерацию панциря. У G. polyedra экдизис вызывается охлаждением клеток до 4 °С в течение 20 мин. При переносе в обычные условия культуры панцирь регенерирует. Этот процесс занимает немного времени: уже через 4 ч на поверхности клетки можно обнаружить элементы панциря, а через 24 ч клетки, "округлившиеся после сбрасывания панциря, принимают характерную для них полиэдрическую форму.
В текальных пластинках есть специальные отверстия — поры для трихоцист. Трихоцисты располагаются по периферии клеток. Число их варьирует от немногих {Gymnodinium simplex) до сотен.
Трихоцисты состоят из узкой области — шейки, прикрепляющейся к теке в месте нахождения округлой поры с приподнимающимся краем, и более широкой части — тела; и шейка, и тело окружены одной мембраной. На поперечном срезе тело имеет квадратные или ромбические очертания и состоит из протеиновых субъединиц, упакованных в форме кубической решетки, шейка же образована пучком спирально закрученных фибрилл. Выбрасываемые при возбуждении через поры в теке трихоцисты значительно длиннее и уже. Они представляют собой белковые нити с периодической исчерченностью под прямым углом к их длинной оси. Есть данные, что примордиальные трихоцисты возникают в центральной части клетки в везикулах — дериватах аппарата Гольджи.
Жгутиковый аппарат. Два жгутика различаются по форме: один «продольный», обычного строения, содержит ак-сонему 9 — 9 + 2, на мембране его во многих случаях в два ряда располагаются волоски; второй — «поперечный» — заключает внутри растянувшейся мембраны, помимо аксонемы, еще исчерченный тяж; мембрана его несет односторонний ряд длинных тонких волосков. Как и у других водорослей, мастигонемы возникают интрацеллюлярно: везикулы, содержащие промастигонемы, в цитоплазме можно видеть часто вблизи ядра и в тесной связи с дикгиосомами и эндоплазма-тическим ретикулумом.
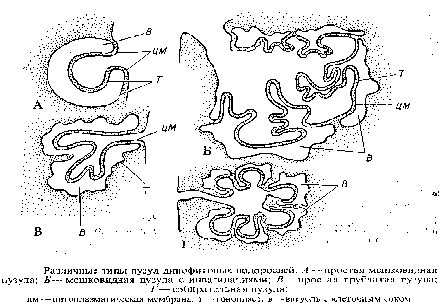
Пузулы. Большинство монадных динофитовых водорослей обладает вместо отсутствующих пульсирующих вакуолей особой органеллой — пузулой. Пузула представляет собой инвагинацию цитоплазматической мембраны в форме мешка или трубки, глубоко вдающуюся внутрь клетки и непосредственно соприкасающуюся с вакуолью клеточного сока. Вакуоль окружает пузулу, стенка которой становится таким образом двух-мембранной. Пузулы изливают свое содержимое наружу через жгутиковые каналы. В случае крупной мешковидной пузулы поверхность контакта между плазмалеммой и тонопластом увеличивается благодаря инвагинациям разветвлений вакуоли внутрь пузулы. То же относится и к трубковидным пузулам. Если приходящее в контакт с вакуолярной системой впячивание плазмалеммы сообща-
ется с наружной средой не непосредственно, а с помощью собирающего пространства, говорят о собирательной пузуле. Расположение в клетке пузырей, собирательных камер, выводковых трубок у разных представителей варьирует. Часто около мешковидной находится собирательная пузула, изливающаяся или в мешковидную, или через свой собственный проток.
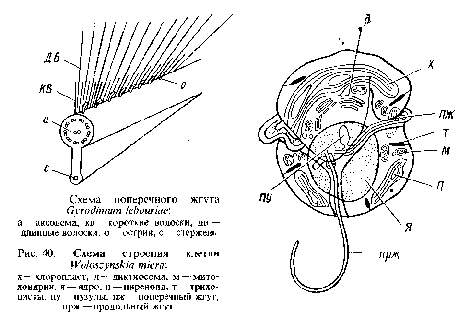
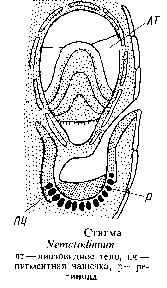
Глазки, расположенные вне хлоропластов у основания жгутиков, также отличаются большим разнообразием: от простейшего глазка в виде массы глобул {Woloszynskia coronata) до сложной светочувствительной органеллы, состоящей из стекловидной, преломляющей свет линзы и пигментного тела, например у рода эритропсис. У нематодиниума стигма тоже состоит из прозрачного линзовидного тела, окружающей его снизу пигментной чашечки и расположенного на дне этой чашечки светочувствительного ретиноидного тела. Линза точно фокусирует свет на дне пигментной чашечки. С физической точки зрения была доказана возможность получения изображения объекта на ретиноидиом слое.
Размножение вегетативное, бесполое, половое. [',,
Отдел Dinophyta делится гта три класса: десмофициевые, динофициевые и бласто-динофициевые. Представители класса дес-мофициевых — монадные, голые или одетые панцирем, преимущественно морские формы со жгутиками, отходящими от переднего конца клетки. Класс динофициевые объединяет монадные, ризоподиальные, паль-меллоидные, коккоидные и нитчатые формы. Монадные клетки дорзивентраль-ные, с поперечной и продольной ложбинками, в которых располагаются жгутики, отходящие в месте пересечения ложбинок на вентральной стороне клетки. Класс бластодинофициевые — искусственная группа, объединяющая паразитные формы.
КЛАСС ДЕСМОФИЦИЕВЫЕ DESMOPHYCEAE -
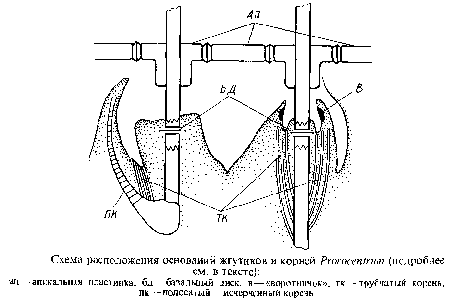
Представителем может служить преимущественно морской род пророцентрум, иногда размножающийся столь обильно, что вызывает цветение прибрежных вод. Клетки более или менее овальные, с боков несколько уплощенные, лишены ложбинок или бороздок. Две главные пластинки тек блюдцевидные, смыкаются вдоль всего края, за исключением их переднего конца, где в одной из них имеется выемка. В апикальной обласги располагается ряд мелких толстых пластинок, довольно прочно соединенных друг с другом так, что апикальная структура имеет тенденцию отделяться как одно целое. От апикальных пластинок отходит и характерный для рода апикальный шип, или зубец,— трубчатая или сдвоенная структура. Главные текальные пластинки гладкие или орнаментированные, перфорированы рядом трихоцистных пор. На переднем конце клетки имеются два жгутика, выходящие через индивидуальные каналы в апикальных пластинках. Один жгут направлен вперед, превышает по длине клетку, имеет заостренный конец и, по-видимому, не несет волосков. Второй жгут, несмотря на то что клетка лишена каких-либо следов поперечной борозды, свойственной монадным клеткам динофициевых водорослей, является по структуре типичным «поперечным» жгутом. Он закручен и внутри свободной мембраны, помимо аксонемы, содержит исчерченный тяж; мембрана его несет односторонний ряд мелких волосков. Основания жгутов в отличие от динофлагеллят параллельны друг другу и располагаются каждое в цитоплазматическом выступе в полости под текой. При вхождении жгутика в цитоплазматический выступ находится толстый трехслойный базальный диск, замыкающий центр аксонемы, под которым периферические дублеты превращаются в триплеты. Оба жгутика различаются по расположению и структуре своих корней. Один жгутик в месте выхода из цитоплазмы имеет «воротничок», и несколько микротрубочек тянутся от основания жгутика к «воротничку». У другого жгута проксимальный конец базального тела заключен в электронно-плотный материал, от которого отходят вперед трубчатый корень и корень с поперечной исчерченностью.
В протопласте находится один желтовато-бурого цвета париетальный сетчатый хлоропласт, окруженный трехмембранной оболочкой и содержащий ламеллы из двух-трех тилакоидов. Пиреноид ограничен растяжением оболочки хлоропласта, матрикс его обычно пересечен несколькими двухтилакоидными ламеллами. Гранулы крахмала откладываются вне хлоропласта. В задней половине клетки расположено ядро, более или менее сферическое, с ядрышком, оболочка ядра с многочисленными порами. Деление ядра такое же, как у других динофлагелля г: ядро становится перфорированным туннелями, которые содержат многочисленные микротрубочки и ориентированы большей частью поперек ядра — под прямым углом к плоскости деления. Тела Гольджи, митохондрии,
мешковидная пузула — как и у других динофитовых водорослей. Предшественники трихоцист находятся вблизи центра клетки, а зрелые трихоцисты — под текой. Размножение осуществляется путем продольного деления клетки, причем дочерние протопласты получают одну половину материнского панциря, а другую выделяют заново.
КЛАСС ДИНОФИЦИЕВЫЕ —DINOPHYCEAE
В зависимости от типа организации таллома класс делится па следующие порядки: перидиниальные, или динофлагелляты -Peridiniales, Dinoflagellatae, динамёби-диальные — Dlnamoebidiales, глеодиниальные — Gloeodiniales, динококкаль-ные — Dinococcales и динотрихальные — Dinotrichales.
Ниже рассматриваются порядки перидиниальные и динококкаль-ные.
Порядок перидиниальные, или динофлагелляты — Peridiniales, Dinoflagellatae
Строение клетки. Это самый обильный представителями порядок динофитовых водорослей. Монадные клетки яйцевидно-шаровидные, более или менее сжатые дорзивентрально, так что при рассматривании с полюсов они имеют эллиптическую или почковидную форму. Нередко клетки несут выросты в виде рогов' или шипов. Большей частью это микроскопически мелкие формы, лишь морская ночесветка видна невооруженным глазом, достигая 2 мм в диаметре. Для тела динофлагеллят чрезвычайно характерны две бороздки. Поперечная бороздка хорошо заметна и обычно полностью опоясывает клетку, отделяя апикальную половину клетки — эпивальву, направленную при движении вперед, от антапикальной — задней половины — гиповальвы. Поперечная бороздка идет или почти горизонтально, как у рода иеридипиум, или чаще несколько по спирали, иногда делая не один, а несколько оборотов, как у рода варновия. Очень редко поперечная борозда опоясывает лишь часть клетки, например у гсмндиннума {Hemidiniwn). Поперечная борозда чаще проходит по середине клетки, но может быть смещена к переднему концу, как у рода амфидиниум, реже к заднему, как у катодини-ума {Katodinium). От положения поперечной борозды зависит высота эпи- и гиповальвы. Эпивальва спереди может быть закруглена или вытянута в длинный рог, как у церациума, гиповальва у церациума также несет один или два рога. Кроме поперечной борозды есть продольная борозда, выраженная менее резко и проходящая перпендикулярно к поперечной по вентральной, несколько сплющенной стороне клетки. Продольная борозда очень немного заходит на эпивальву и часто ограничивается одной гиповальвой — антапикальной стороной клетки.
Однако у рода гоннаулакс продольная борозда простирается от переднего до заднего конца клетки. У некоторых морских перидиней продольная борозда не прямая, а изогнутая или спирально извитая.
Два жгутика динофлагеллят выходят на вентральной стороне клетки через индивидуальные отверстия в текальных пластинках. Поперечный лентовидный жгут опоясывает клетку и скрыт в поперечной борозде, лишь дистальный конец его выступает наружу. Жгутиковая мембрана поперечного жгута растянута и, как правило, заключает узкий негибкий тяж, имеющий правильную поперечную полоса-тость. Этот тяж находится в состоянии натяжения, что, по-видимому, и удерживает поперечный жгут в борозде. Гораздо более длинная аксонема идет по спирали вокруг тяжа. Поперечный лентовидный жгут несет односторонне расположенные волоски одинаковой длины {Gonyaulax polyedra, Gymnodinium breve и др.) или же волоски могут различаться по длине. Например, поперечный жгут Gyrodinium iebouriae несет длинные и короткие волоски и еще третий гип придатков в виде сплошных, изогнутых по направлению к кончику жгута шипов. Все три типа отростков располагаются вдоль одного спирального ряда. На самом кончике жгута волоски и острия отсутствуют.
Продольный жгут значительно длиннее продольной борозды, выступает из нее и направлен по движению клетки назад. Тонкие волоски располагаются на нем в два ряда {Gyrodinium Iebouriae, Woloszynskia micra, Oxyrrhis marina). Жгутиковые волоски образуются в расширенной перинуклеарной полости. Расположение жгутиковых корней очень варьирует.
На рис. 40 приведена схема строения монадной клетки динофициевых водорослей.
Размножение динофлагеллят вегетативное, бесполое и половое. Вегетативное размножение осуществляется делением клетки пополам, например у гимнодиниума, цератиума, перидиниума, го-ниаулакса.
Клеточный покров динофлагеллят—тека — интенсивно изучался у взрослых, более старых клеток. Гораздо меньше сведений о покровах во время и вскоре после деления клетки. У панцирных динофлагеллят можно различить два разных типа деления клетки. При одном типе, характерном для видов цератиум, гониаулакс старый панцирь материнской клетки распределяется на две дочерние клетки, а недостающие половинки панциря вырабатываются заново. При другом типе деления клетки, характерном для рода перидиниум {Peridinium cinctum), дочерние клетки образуют новый панцирь
полностью, а старый при этом погибает. Г. Дюр исследовал оба типа: первый на примере G, polyedra, второй — у P. cinctum.
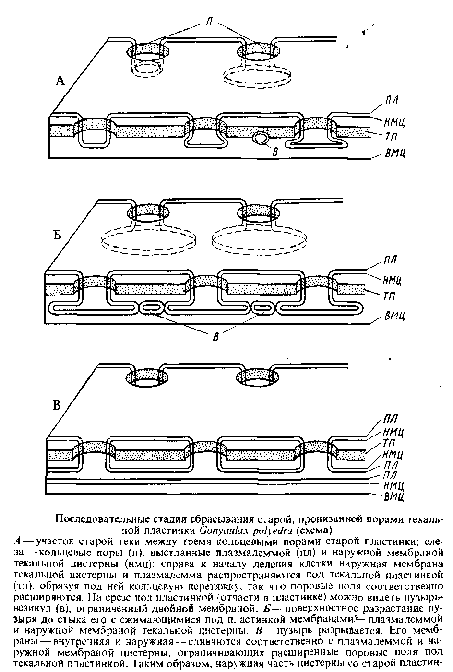
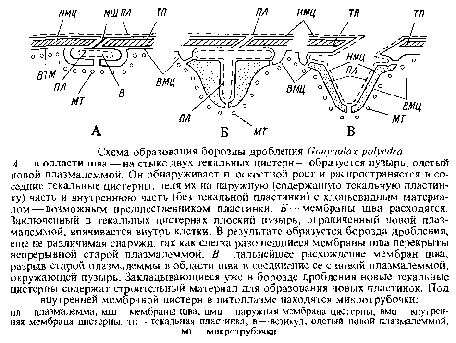
У G. polyedra после деления ядра появляется борозда дробления. Эта борозда закладывается почти одновременно по всей теке, следуя линии швов, мест соединения — стыка мембран текальных цистерн. Мембраны шва разъединяются на две мембраны соседних лекальных цистерн. Возникающий в этой области пузырь, одетый мембраной, распространяется в плоскостном направлении и внедряется в соседние текальные цистерны под находящиеся там пластинки. Этим текальные цистерны расщепляются на две части: наружную с текальными пластинами, и внутреннюю, содержащую лишь зернистый строительный материал для новых пластин. Плоские пузыри, заключенные в текальные мембраны, затем впячиваются по направлению к центру клетки под несколько перекрывающими друг друга пластинами панциря и сливаются. Так возникает борозда дробления, снаружи еще не различимая. Одновременно внутрь клетки смещаются и внутренние, нижележащие части текальных цистерн. Таким образом, в борозде дробления под плазмалеммой уже с самого начала находятся уплощенные пузыри — цистерны, содержащие строительный материал для новых пластинок панциря. Под ними находится слой цитоплазмы с микротрубочками. Позднее борозда прорывается наружу: мембраны шва расходятся, внутренняя новая плазмалемма пузыря соединяется с наружной старой плазмалеммой и обе дочерние клетки отклоняются друг от друга.
В результате деления возникают две дочерние клетки, теки которых состоят на одну половину из старых текальных пузырей с пластинами, а на другую — из текальных пузырей, еще не содержащих пластин. Возможно также, что обе половины теки уже с пластинами, но пластины различаются по степени развития скульптуры, однако эта разница между двумя половинками теки в молодых дочерних клетках очень быстро исчезает, и уже вскоре после деления почти все клетки Gonyaulax polyedra обнаруживают однородно скульптурированный панцирь. Такая однородная структура панциря молодых клеток достигается в результате обновления старой половины панциря, унаследованной от материнской клетки.
Еще во время деления клетки в текальных цистернах и отчасти в пластинах появляются везикулы с двойной стенкой. При увеличении этих везикул в направлении, параллельном поверхности, как и при образовании борозды дробления, текальные цистерны расщепляются на наружную часть, содержащую пластинку, и внутреннюю, возможно, с остатками старой пластинки. При разрыве пузыря-везикула у расширенных краев кольцевых пор, выстланных плазмалеммой и наружной мембраной цистерны, внутренняя мембрана пузыря соединяется с плазмалеммой, а наружная --с наружной мембраной текальной цистерны. Наружная часть текальной цистерны, содержащая старую пластинку, пронизанную порами, изолируется и сбрасывается, а во внутренней части возникает новая пластинка.
Таким образом, старые пластинки панциря сбрасываются в результате перешнуровки текальных пузырей в плоскости, параллельной поверхности. Внутренняя часть перешнурованных текальных пузырей, возможно, содержит еще старый материал для образования новых лластинок. Эти пластинки не будут отличаться от новых пластинок, возникающих в пузырях, выстилающих борозду дробления, т. е. в половинке клетки, образующей панцирь заново.
У Peridinium cinctum клетка делится внутри старого панциря, текальные пластинки которого отчасти растворяются. Дочерние клетки удерживаются некоторое время вместе внутри разрушающегося панциря, заключенные в общий фиброзный слой; после его разрыва клетки освобождаются. Еще внутри старого панциря в дочерних клепках начинается отложение текальных пластинок. Ультратонкие срезы через делящиеся клетки позволили проследить возникновение новой плазмалеммы дочерних клеток. Под внутренней мембраной текальных цистерн старого панциря появляются плоские везикулы, которые сливаются с образованием наружной и внутренней мембран. Внутренняя мембрана пузыря становится плазмалеммой дочерних клеток. Из содержимого пузыря, возможно, возникает фиброзный слой. Наружная мембрана везикула остается с внутренней мембраной текальных цистерн, старыми текальными пластинками, наружной мембраной текальных цистерн и старой плазмалеммой.
У Woloszynskia apiculata при бесполом размножении клетки теряют подвижность, округляются и превращаются в зооспорангии. При этом под текой выделяется тонкая целлюлозная оболочка собственно зооспорангия. Содержимое зооспорангия делится на две, четыре, восемь клеток, которые освобождаются при растворении целлюлозной стенки собственно зооспорангия и разрыве и сбрасывании теки исходной материнской клетки.
На протяжении двух последних десятилетий у многих дино-флагеллят рядом ученых обнаружено половое размножение, ранее известное лишь у одного из видов гленодиниума. Успех этих исследований в большой степени обусловлен развитием методов культивирования на искусственных питательных средах и накоплением данных о том, что половой процесс у многих водорослей индуцируется недостатком в среде азота. Это оказалось справедливым и для многих динофлагеллят. Так, при помещении в среду, свободную от азота, или в старых культурах, где уровень азота низок, половой процесс наблюдался у ряда видов Peridinium Woloszynskia apiculata, Glenodinium lubiensiforme, Gonyaulax moniliata, Crypthecodinium cohnii, Protogonyaulax catenella и др. Сливаются голые гаметы одинакового размера, г, е. имеет место изогамия, и зиготы у P. cinctum, P. willei, P. Umbatum, Gymnodinium pseudopalustre, Crypthecodinium cohnii возникают в пределах одного клона.
Woloszynskia apiculata изогамна, но гетероталлична, у этого вида наблюдалось образование групп. Гаметы сливаются своими боками. Напротив, у видов рода цератиум {Ceratium horridum, С. cornutum) копуляция гамет постоянно анизогамная, и сам процесс слияния иной: мужская гамета входит в женскую через отверстие поблизости от места прикрепления жгутиков.
Отличительной чертой всех до сих по"р изученных в этом отношении динофлагеллят является долго продолжающийся рост планозиготы. Так, планозиготы Gymnodinium pseudopalustre, прежде чем превратиться в покоящуюся зиготу, растут 11 дней при 15'' С или 8 дней при 21 ° С. У Woloszynskia apiculata планозиготы также сильно вырастают и остаются подвижными в течение 8—10 или 3—4 дней при температурах 15 и 21° С соответственно. Планозиготы передвигаются с помощью двух сохраняющихся задних жгутиков гамет, в некоторых случаях остаются и поперечные жгутики, по крайней мере один из них. Динофлагелляты с «двойными» задними жгутиками описывались более 100 лет тому назад и в материале из природы: по-видимому, в этих случаях речь идет о планозиготах, которые нередко таким образом возникают и в природных условиях, а не только в культурах. После длительного движения, сопровождающегося ростом, планозиготы превращаются в неподвижные толстостенные гипнозиготы.
Г. Дюр исследовал строение оболочек гипноспор в электронном микроскопе. Наружная оболочка похожа на панцирь вегетативных клеток, отличаясь от него большей толщиной пластинок и более широкими швами. Образование средней и внутренней оболочек, по-видимому, сходно с образованием теки, т. е. они состоят из пластинок, которые возникают в текальных цистернах, однако швы отсутствуют. Это заставляет предположить, что цистерны, в которых образуются пластинки мезоспория и соответственно эндоспория, сливаются с образованием непрерывных наружной и внутренней мембран: затем между ними возникают мезоспорий и эндоспорий.
Через некоторый период покоя зиготы прорастают: так, отдельные зиготы Gymnodinium pseudopalustre и Woloszynskia apiculata способны прорастать через несколько недель на свету при 15" С. Однако более обильное и синхронизированное прорастание бывает при охлаждении зигот по крайней мере в течение 4 недель при 6 С для Woloszynskia и при 3е С для Gymnodinium и последующем переносе их на свет в условия более высокой температуры. При прорастании зигот всех изученных до сих пор динофлагеллят происходит мейоз, и таким образом динофлагелляты -—гаплонты, т. е. их ядра в вегетативных клетках имеют гаплоидный набор хромосом.
* * *
Упомянутые выше роды динофлагеллят распределяются по многим семействам. Кроме них можно назвать еще род ночесветка {Noctiluca) и ряд родов, объединяемых в семейство динофизовые, заметно отличающиеся от вышеописанных динофлагеллят.
Род ночесветка — крупная, распространенная по всему свету морская бесцветная голозойная динофлагеллята. Клетка сферическая, достигающая 1 - 2 мм в диаметре, покрыта снаружи полиэдрическими, одетыми мембраной текальными везикулами, снабженными трихоцистными порами. Изнутри везикулы выстланы рядами микротрубочек, тянущихся параллельно поверхности клетки. Цитоплазма образует гонкий периферический слой, от которого отходят тонкие тяжи, пересекающие крупные вакуоли с клеточным соком, Постенная цитоплазма содержит многочисленные липидные везикулы; некоторые из них, возможно, являются «микроисточниками» биолюминесценции. На одной стороне сферической клетки имеется глотка, в которой расположен один короткий жгутик. Из этой части клетки образуется крупное поперечно исчерченное подвижное щупальце, функция которого — улавливать необходимые для питания организмы в свой слизистый эксудат и увлекать их в глотку, где они заключаются в пищеварительные вакуоли. В цитоплазме находятся ядро и крупные митохондрии, трихоцисты. По соседству с местом прикрепления щупальца расположены пучки исчерченных волокон, возможно, контролирующих его движение.
Наряду с делением — наиболее обычным способом размножения у ночесветки описаны: образование многих зооспор, похожих на гимнодиниум, и половой процесс в.виде слияния од-ножгутиковых изогамет, возникающих внутри материнской клетки в числе 1024. Есть данные, что перед образованием гамет происходит мейоз, за которым следуют многие синхронные митозы. Если эти данные подтвердятся, то ночесветка будет представлять исключение среди других динофлагеллят, у которых редукция зи-готическая.
Семейство динофизовые включает главным образом обитателей теплых тропических морей. Клетки дорзивентральные, сжатые с боков, снабжены, как и другие динофлагелляты, бороздками, причем поперечная борозда сильно смещена вперед. Таким образом, эпивальва небольшая и жгутики отходят в месте пересечения борозд также недалеко от переднего конца клетки. Клетка одета текой, в везикулах которой находятся толстые целлюлозные текальные пластинки. Текальные пузыри с пластинками, соприкасаясь краями, образуют панцирь, который здесь в отличие от других динофлагеллят разделен на две боковые половинки, соединенные продольным швом. Края как продольной, гак и поперечной борозд несут крыловидные выросты. Размножение—продольным делением клетки пополам в плоскости сагиттального шва: каждая дочерняя клетка получает половинку панциря материнской клетки, а недостающую строит заново.
Род динофнзис. Края поперечной борозды направлены вперед, образуя воронки, скрывающие небольшую эпивальву. Продольная борозда несет крылья, несколько различающиеся по размерам и форме. Некоторые виды встречаются и в холодных морях {D. norvegica) и лиманах.
Род орнитоцеркус характеризуется эпивальной, редуцированной до маленьких пластинок, соединенных продольным швом. Крыловидные выросты очень сильно развиты как у продольной, так и у поперечной борозды.
Перилинеи распространены в планктоне как морей, так и пресных вод. Они занимают второе место как продуценты органического вещества в морс.
Наряду с пользой псридинеи могут приносить и вред. Среди них имеются паразиты, некоторые из них нападают на личинки устриц. Есть виды, вырабатывающие яд, смертельный для рыб. Кроме того, разлагаясь после своего массового развития, они могут отравлять воду на многие километры вредными продуктами распада, вызывая замор рыб и других водных животных.
Порядок динококкальные — Dinococcales
Объединяет коккоидные формы — неподвижные, одетые оболочкой клетки с типичными для динофитовых водорослей внутренним строением. Вегетативное деление отсутствует, размножение зооспорами, автоспорами. В ряде случаев замена зооспор автоспорами может быть прослежена у разных видов в пределах одного и того же рода.
У рода тетрадиииум клетки имеют форму тетраэдра, углы которого вытянуты в простые или двойные шипы.
При зооспорообразовании протопласт слегка сокращается и делится пополам, после чего у продуктов деления формируются бороздки, характерные для монадных клеток динофициевых водорослей, появляются стигмы, В конце концов в растянувшейся оболочке материнской клетки можно видеть две гимнодиниумподобные зооспоры, позднее освобождающиеся через разрыв оболочки.
Род мистодиниум, встречающийся в болотах, характеризуется веретеновидными или полулунными клетками. У одних видов размножение зооспорами: содержимое клетки отстает от оболочки и еще до деления принимает форму динофлагелляты, т.е. развивает систему бороздок, появляется стигма; однако жгутики на этой стадии не образуются. Такая клетка делится на две гимнодиниумподобные зооспоры, вырабатывающие уже жгутики и покидающие оболочку материнской клетки. После краткого периода движения зооспоры останавливаются, бороздки пропадают, протопласт вытягивается и выделяется новая оболочка. У Cystodinium lunare наблюдается редукция подвижной стадии: при размножении протопласт сокращается, но, не вырабатывая бороздок и стигмы, претерпевает деление; продукты деления, также, не образуя бороздок и стигмы, несколько вытягиваются, принимают очертания материнской клетки, одеваются собственной оболочкой и освобождаются в виде мелких клеток цистодиниума. Таким образом, здесь вместо образования зооспор происходит автоспорообразование.

Нравится материал? Поддержи автора!
Ещё документы из категории биология:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ