Поведінка самців щурів лінії Вістар при різних світлових режимах
 Зміст
Зміст
Вступ
1. Біоритми, як загальні властивості живого
1.1 Структурні елементи біоритма
1.2 Класифікація біоритмів
1.3 Добові біологічні ритми
2. Поведінкові реакції тварин і методи їх вивчення
2.1 Форми поведінки тварин
2.2 “Відкрите поле”, як один із методів вивчення оведінкових реакцій тварин
3. Методи та матеріали дослідження.
3.1 Методика вироблення штучного циркадного біоритму у самців щурів лінії Вістар
3.2 Методика проведення тесту “Відкрите поле”
4. Поведінкові реакції щурів які утримувались у різних вітлових режимах
4.1 Поведінкові реакції самців щурів контрольної групи
4.2 Поведінкові реакції самців щурів Д-Н групи
4.3 Поведінкові реакції самців щурів Д-групи
Висновки
Література
Вступ
В основі життєдіяльності живого організму лежать ритмічність і рухомість його фізіологічних, біохімічних та біофізичних функцій. Це енергетично вигідна і найбільш оптимальна форма існування. Узгодженність у часі і просторі всіх життєзабезпечуючих систем зумовлює нормальне функціонування організму. Напруженність його функціонування, в свою чергу, свідчить про інтенсивність впливу зовнішніх і внутрішніх факторів середовища. Всі ці явища сьогодні розуміються як біоритми - ритми життя.
Одним з основних критеріїв резистентності організма людини є стан системи добових ритмів організма. Добовий ритм фізіологічних процесів і поведінки є у всіх живих організмів. Він погоджений з добовою періодичністю зміни метеорологічних факторів. Знання динаміки добової ритміки фізіологічних показників є цінним діагностичним (оцінка причинно-наслідкових факторів виникнення хвороби) і прогностичним (компенсаторні можливості організма, прогноз розвитку патологічного процеса) прийомом, особливо в процесі пристосування до природних умов існування.
Відомо, що світло, особливо сонячне, чинить глибокий вплив на обмінні процеси організма. Тому, саме за цією причиною одним з найсильніших зовнішніх синхронізаторів біоритмів є зміна освітлення (Pittendrigh, 1965; Aschoff, 1968, 1969; Wever, 1968, 1969, 1970). Перебудову фізіологічних ритмів при експериментальній зміні освітлення спостерігали такі вчені, як О.П. Щербаков і Г.М. Черкович. В останні часи ця проблема привертає до себе все більше і більше уваги в зв’язку з частою позмінною працею населення, постійним відвідуванням молоддю різноманітних нічних клубів та інтернет-кафе.
З клінічних досліджень виявлено, що нічна праця чинить негативний вплив на системи травлення і кровообігу. Pierach (1955) У людей, які працюють в нічну зміну, в порівнянні з іншим населенням, набагато частіше
проявляються хвороби шлунково-кишечного тракту - від гастрита до язви. При цьому язва шлунку і дванадцятипалої кишки у працюючих вночі важче піддається лікуванню, частіше спостерігаються рецидиви, що ведуть до інвалідності. [4] Порушення серцево-судинної функції пов’язані з невідповідністю між необхідністю посилення функціонального напруження системи кровообігу і підвищенням тонусу парасимпатичного відділа нервової системи (падіння систолічного тиску, уповільнення пульсу і т.д.) в нічні години. Нічна праця призводить до зміни добового біоритму серцево-судинної діяльності - ритм приймає монотонний характер. А це в свою чергу, призводить до виникнення атеросклерозу (експериментальні роботи С.І. Серова, Г.М. Цветаєвої, 1979), частих приступах стенокардії, зниження толерантності до фізичного навантаження. У людей, працюючих в нічну зміну, порушується обмін речовин. відбувається ніби розмежування між доставкою в організм енергетичних з’єднань і швидкістю їх утилізації. І внаслідок цього в крові накопичуються недоокислені продукти обміну, спостерігається її закислення. [2] Нічна праця зменшує циркадну організацію екскрекції натрію зі слиною, що відбиває ритмічне виділення металокортикоїдних гормонів. Т.Д. Семенова (1971)
Світло в першу чергу впливає на функціонування зорового аналізатора. Так, під час перебування криз в режимі цілодобового освітлення у них збільшився офтальмотонус майже до 30 мм. рт. ст. Т.А. Красновид (1972)
Вночі зменшується координація рухів, лабільність нервово-м’язового апарату, сила м’язів і їх статистична витривалість. Праця, що викликає однакове підвищення ЧСС, вночі сприймається як більш важка в порівнянні з денним відчуттям. Витривалість, особливо до статичних зусиль, якість виконання роботи в нічні години помітно знижується. Людина, що працює передусім вночі, часто буває і більш роздратованою.
Вплив різних режимів освітлення на функціонування організму інтенсивно вивчався ще на початку XX століття, а зараз практично не розглядається. В сучасних роботах недостатньо клінічних та експериментальних даних.
Виходячи з актуальності теми, метою проведення нашого дослідження було дослідити поведінку самців щурів лінії Вістар при різних світлових режимах за допомогою методики “Відкрите поле”.
Завданнями роботи є:
проаналізувати науково-методичну літературу з даної теми дослідження;
визначити показники рухової активності, грумінгу, рерінгу, дефікації і інших складових поведінки щурів і порівняти їх з літературними даними;
дослідити можливість використання даної теми дослідження на уроках біології та в позакласній діяльності вчителя.
Предметом дослідження є поведінка самців щурів лінії Вістар при різних світлових режимах.
Об’єкт дослідження: самці щурів, які знаходились в різних світлових режимах утримання.
1. Біоритми, як загальні властивості живого
Життєдіяльність різноманітних істот, незалежно від складності їх організації, протікає в незакінченній зміні станів, незупинної адаптації до навколишнього середовища за рахунок пластичності і рухомості властивих їм функціональних можливостей. Але подібна перебудова організму і погодження його систем з постійно змінюючимися умовами навколишнього середовища можливі лише при наявності безперервних коливальних процесів всередині організму. Причини цього в тому, що". адаптація, являючись однією з фундаментальних властивостей біосистеми, тісно пов’язана з її іншою фундаментальною властивістю - циклічностю. ”. Баєвский Р.М. (1979)
Основою гомеостаза і гомеокінеза - властивості організма зберігати постійність внутрішнього середовища і в той же час змінювати функціональний стан організму у відповідності з умовами навколишнього середовища - є ритмічна активність всіх систем, їх гармонійне пропорційне поєднання. Ритмічні коливання процесів являють собою загальну характеристику активності біологічних систем і основу їх регуляції і цілісності. [1]
Ритмічність і діапазон коливання фізіологічних, біохімічних, біофізичних і імунологічних процесів лежать в основі життєдіяльності живого організму. Періодичність функцій - енергетично вигідна і найбільш оптимальна форма життєдіяльності. Погодженість у часі і просторі всіх життєзабезпечуючих систем зумовлює нормальне функціонування організму. Напруженість його існування, в свою чергу, свідчить про інтенсивність впливу зовнішніх і внутрішніх факторів середовища. Всі ці явища сьогодні розуміються як біоритми - ритми життя. За образним висловлюванням Н.Я. Перна “життя - є складна тканина, що складається з безкінечних різноманітних ритмів”.
щур біоритм поведінкова реакція
С.И. Степанова (1977) стверджує, що життя - це ритм, втрата якого рівносильна смерті. Таким чином, виникнення життя йшло в напрямку ускладнення і удосконалення просторової і часової організації.
Біоритми - це послідовне чергування процесів напруги і розслаблення (Bunning, 1961, 1973), повторення однієї і тієї ж послідовності подій у фізіології координації часу (Hal berg, 1964, 1977). Вони являються невід’ємною властивістю живих систем і складають основу їх організації, незалежно від періодичності зовнішніх змін (Pittendrigh, 1964). За A. Sollberger (1965), це регулярні і якісні зміни біологічних процесів на всіх рівнях організації живого. B. Goodwin (1966), наголошує, що упорядкованість біосистем, існує не лише в просторі (морфологія організмів), але і в часі, про що свідчить погодженість їх поведінки.
Біоритми проявляються в багаточисленних адаптивних реакціях на періодичні зміни факторів навколишнього середовища. Вони являють собою цілий комплекс цитологічних, фізіологічних, поведінкових і інших реакцій, направлених на підтримку оптимальної послідовності фаз життєдіяльності в умовах коливання впливів середовища. Чим точніше біологічні реакції організма приурочені до окремого часу доби чи року, тим вище їх адаптація до умов існування. Істоти, які в ході еволюції виробили різні форми добової активності, змогли більш повніше освоїти природні міста середовища існування. Наявність в біогеоценозах суспільств, активність яких має періодичність, сприяє досконалішому використанню організмами енергетичних ресурсів по всій території. Завдяки різному ритму життя різні види можуть існувати на одній території, зберігаючи при цьому видову ізоляцію. В залежності від кліматичних умов змінюється характер біоритмів активності організмів. Різниця в протіканні активних фаз біоритмів однієї популяції спеціалізують групи по тим чи іншим “часовим” нішам і сприяють їх ізоляції. Використання різними видами різних “часових” ніш в межах однієї екологічної ніші являє собою еволюційний процес адаптативної радіації. Разом з тим функціонально-динамічні властивості організма відрізняються окремою стійкістю, константністю і, навіть, консервативністю, що являється проявом індивідуальних особливостей біоритміки і реактивності. Тому механізми адаптації і збереження добового динамічного стереотипу тісно пов’язані з стійкістю біоритмічного гомеостазу
(Василевський Н.Н., 1973; Баевський Р.М., 1977; Моісеєва Н.І., 1978; 1980).
Отже, біоритми - це функціональні системи організма, що забезпечують гомеостаз внутрішнього середовища, і адаптаційні реакції за відношенням до факторів навколишнього середовища побудовані так, що їх регулювання направлене на збереження цілісності системи, підтримку її стійкості і збереження рівноваги як єдиної функціональної системи на основі законів симетрії і гармонії. Завдяки наявності власних коливальних систем організм має здатність сприймати ритм навколишнього середовища та узгоджувати з ними власні ритми, проводячи настрій і сонастрій всіх підсистем у відповідності зі своїми потребами. При цьому зовнішні часові фактори не індукують ритмів, що перебігають в організмі, а лише впливають на них, - “захоплюючи”, модулюючи і т.д. Ось чому, біологічний годинник - “механізм часової організації функціональних процесів”, закріплено на кожному структурному рівні організації організму.
1.1 Структурні елементи біоритма
Життя організма складається з безперервних процесів життєдіяльності протягом доби, тому найбільш важливою є циркадна ритміка змін показників і параметрів стану організма. Біоритміка живого багатозначна - вона складається з найрізноманітніших ритмічних коливань, протяжністю від долей секунди до тривалих багатоденних чи багатомісячних змін (Сисуєв В.М., 1981; Hal berg F., 1969; Sinz R., 1978). Але, не дивлячись на цей широкий діапазон коливань, вони всі мають окремі елементи, схожі у всіх ритмічних типах.
Біоритм відображає зміни якого-небудь показника чи властивості протягом окремого часу, тому при графічному зображенні ритма на одній осі координат відкладають величину досліджуваного показника, а на іншій осі - інтервал часу проведення вимірів. Зміни функціональних показників можна умовно уявити у вигляді синусоїди і на ній виділити такі структурні елементи ритма: період, амплітуду, фазу, акрофазу. Перпіод - проміжок часу між двома повторюючимися вихідними точками ритмічної кривої, наприклад, між двома підйомами чи пониженнями, що називаються фазами ритма. Амплітуда - величина варіації досліджуваного показника чи, інакше, половина величини періодичної зміни в даний момент, взяту як різницю між максимальним відхиленням і середнім значенням. Часто її виражають в процентах від всіх значень амплітуд за час спостереження ритму і називають мезором, який можна приблизно визначити на графіку, провівши пряму лінію по середині відстані між найбільшими значеннями амплітуд кожної фази ритма. Час максимального відхилення досліджуваного показника протягом періоду ритма називається акрофазою.
Одним з найважливіших елементів біоритма є його базисний рівень, вимірюємий як мода ряда найменших значень абсолютних величин показника. Він являє собою той рівень функціонального показника, на якому виявляються коливання в сторону збільшення чи зменшення амплітуд і по всій величині базисний рівень може бути низьким, середнім чи високим. Ця властивість біоритмів лежить в основі їх класифікації. Базисний рівень звичайно розраховують за довгий проміжок спостереження ритміки (тиждень, місяць) і при його визначенні не приймають до уваги різкі пониження показника, так як він виражає моду ряда мінімальних значень показника. Аналіз робіт з біоритмів доводить, що зміна функціональних показників у різних організмів може бути схожа за періодом, частотою, фазою, але протікати на різному базисному рівні. Останній, для значень якого-небудь показника, наприклад, вмісту гормонів, цукру, мікроелементів в крові, сечі, слині, величина артеріального тиску і інші показники, являють собою досить стійку величину для досліджуваного, що зберігається довгий час майже незмінною, і її зміна може спостерігатися лише в момент проходження важливих фаз в онтогенезі (пубертальний період, клімакс і т.д.). У випадку хвороби чи якихось інших різких змін стану органі (стресова реакція) базисний рівень може різко змінюватись, що зразу наголошує на порушенні в стані функціональних систем, регуляторної функції нервової системи і зміні внаслідок цього в системі гомеостаза.
Важливою додатковою характеристикою біоритма є також розмах коливань, т. б. величина тих меж, в межах яких відбуваються коливання досліджуваних показників, чи, говорячи іншими словами, варіабельність значень амплітуд досліджуваних показників за відношенням до мезора. Цей показник показник характеризує пластичність досліджуваних функцій організма, степінь його адаптованості за даним параметром, варіабельність параметрів біоритма.
Проведені дослідження доводять, що високі функціональні можливості організма: стійкість до дії інфекційних агентів, виражена адаптація при тривалих перельотах, висока працездатність і т.д., спостерігаються у людей з відносно високим базисним рівнем, великим розмахом коливань, стійкою часовою структурою біоритма і відносною постійністю акрофаз (Моісеєва Н.І., Сисуєв В.М., 1981). В той же час встановлено, що найбільш легко пристосовуються до змінної праці ті люди, у яких відмічені низькі значення амплітуд біоритмів окремих показників (Rein berg A. et al., 1978).
Для точної оцінки, опису і аналізу біоритмів введені також і інші показники, наприклад, коефіцієнт синхронізації функцій, показники добової активності (Баевський Р.М., 1976), коефіцієнт степені організованості добового ритма (Моісеєва Н.І., Сисуєв В.М., 1981) і ряд інших показників оцінки біоритмів, що мають спеціальне значення (Halberg F. et al., 1977). Всі ці показники є похідними від основних параметрів ритма.
1.2 Класифікація біоритмів
Біологічні ритми охоплюють широкий діапазон періодів - від долей секунд на молекулярному рівні, до багатьох років - на надорганізмовому. З підвищенням рівня структурної організації матерії подовжується період набутих нею коливань. Ускладнення функціональної організації системи веде до створення ієрархічної черговості часових рівнів.
Існують класифікації біоритмів, що грунтуються на їх функціональній значимості, на рівні організації біологічної системи, рівні гомеостатичних механізмів. Найбільше розповсюдження має поділ біоритмів в залежності від їх частоти (періоду).
Найбільш повна класифікація частоти біологічних ритмів дана в праці Ашоффа, в якій виділено п’ять класів біоритмів:
ритми високої частоти - від долей секунди до 30 хвилин (осцилляції на молекулярному рівні, ритми електроенцефалограмми, скорочення серця, дихання, перистальтика кишечника);
ритми середньої частоти - від 30 хвилин до 28 годин, включаючи ультрадіальні (до 20 год) і циркадні (20 - 28 год);
мезоритми, до яких відносяться інфрадіальні (28год - 6 днів) і циркасептальні (біля 7 днів);
макроритми з періодом від 20 днів до 1 року;
Мегаритми з періодом в десятки і багато десятків років.
Існують також інші класифікації [9], згідно яким виділяють 6 головних ритмів:
добові, зумовлені 24-годинною сонячною активністю, але, проте, часто вони стають навколодобовими, чи циркадними;
лунні ритми, пов’язані з добою місяця, продовженість якої дорівнює 24години 50 хвилин і визначаються періодом зміни положення Місяця за відношенням до Землі;
ритми зірок, що визначаються періодом доби зірок, т. б. оборотом Землі навколо власної осі і періодичними (23 год 56 хв) змінами її положення за відношенням до зірок;
півмісяцеві і місяцеві ритми (протягом місяцевого періода два рази змінюється амплітуда приливів, якщо вектори сил тяжіння Місяця і Сонця співпадають, то приливи максимальні, якщо вони знаходяться під кутом 90°, то мінімальні);
річні ритми;
багаторічні ритми (біля 11 років триває період коливання сонячної активності, зумовлених максимальними і мінімальними фазами сонячних плям).
Відомі короткоперіодні ритми активності тварин. До цього класу відносять різні періодичні повторення якої-небудь форми поведінки в різний період добового цикла з частотою від 20 хв до 6 год. Виключення складають як циркадні, так і високочастотні ритми в нервовій системі і в м’язах, пов’язані з інтеграцією рухів. У більшості випадків короткоперіодичні ритми рухомості пов’язані з потребами їжі.
Данні прижиттєвої кількісної мікроскопії, мікрокінозйомки і культивування тканин показали, що існують клітинні ритми великих діапазонів. Експериментально знайдено наявність швидких (до 100 мкс) коливань окремих мікроструктур і встановлений діапазон періодів їх фонових ритмів в одній і тій же живій клітині.
Складений перелік ряду біологічних ритмів в порядку зменшення їх частоти. Найбільш високою частотою відрізняються біотоки 60 - 100 коливань в 1 секунді, потім йдуть коливання війок мерехливого епітелія, хвилі ЕЕГ, пульс, дихання, перистальтика кишечника. Протяжність періода цих ритмів складає долі хвилини. Далі йдуть ритми, періоди яких досягають декількох годин: коливання числа тромбоцитів і рівня адреналіна в крові.12-годинні ритми характерні для вмісту глікогена в печінці, числа ретикульоцитів, а також вмісту білірубіна і жиру в крові. [16]
Періодичні зміни біологічних процесів (як кількісні, так і якісні), що відбуваються на різних рівнях організації живого - будь то молекулярно-генетичний, клітинний, тканевий, органний чи, навіть, популяційно-біосферний рівень, - вчені охрестили біоритмами і в відповідності з виконуваною функцією розділили їх на фізіологічні (т. б. робочі цикли окремих систем організма) і екологічні, чи адаптивні, пов’язані з пристосуванням до періодичних впливів зовнішнього середовища.
Згідно класифікації відомого хронобіолога Ф. Хальберга всі ритмічні процеси організма у відповідності до власної довжини діляться на декілька груп: ритми високої частоти з періодом до 30 хвилин, ритми середньої частоти з періодом від 30 хвилин до 6 днів і ритми низької частоти з періодом від 6 днів до 1 року. До першої групи відносяться ритми дихання, роботи серця, електричних явищ в головному мозку і періодичні коливання в ланцюгу біохімічних реакцій. До другої - зміна сну і бадьорості, активності і відпочинку, добові зміни обміну речовин. третя група об’єднує тижневі, місяцеві і річні ритми, що включають цикли екскреції гормонів, сезонні зміни протікання біохімічних реакцій і довготривалі часові змінм працездатності.
Система ритмів ієрархічна. Є ритми ведучі і відомі. До ведучих відносяться ритми центральної нервової системи, ендокринних залоз, до відомих - ритми температури, пульса, обміна. Фізіологічні і екологічні ритми - циклічні коливання в різних системах організма - несуть постійний характер і майже не залежать від температури і складу середовища. Є здогадки, що регуляція фізіологічних ритмів здійснюється гіпоталамо-гіпофізарною системою.
Екологічні ритми залежать від циклічних змін навколишнього середовища, завдяки чому в окремі інтервали часу організм може бути готовий до сприйняття сигнала (так називаємий час потенційної готовності). Вони відчувають більший вплив факторів середовища (добові, приливні, місяцеві, річні ритми).
Біоритми в природі складаються з ендогенного ритму і реакції на зміни середовища. При порушенні природного ритму середовища чи власних центрів синхронізації біоритми різних фізіологічних функцій втрачають синхронність.
Добові ритми пов’язані з одним оборотом Землі навколо власної осі. При цьому протягом доби закономірно змінюються освітлюваність, температура, вологість, напруженість електричного і магнітного полів Землі і, навіть, інтенсивність потока сонячних частинок, що має життєво важливе значення для різних мешканців нашої планети. Добова періодичність властива всім без винятку організмам і внутрішньоклітинним процесам, а також життєвим процесам взагалі. У людини, наприклад, описані більш 400 ритмопроявляючихся функцій, що повторюються протягом доби і погоджених один з одним. Чіткість погодження не абсолютна, і добові максимуми і мінімуми різних функціональних показників на 24-годинній шкалі розділені відносно постійними часовими інтервалами, чи фазовими кутами, тому таку незмінну протягом багатьох діб погодженість називають фазою. Циркадні ритми знайдені також і у окремих клітин багатоклітинного організму. У людини і тварин центри синхронізації добових ритмів, чи пейсмекери (водії ритмів), що керують ритмами клітин, органів і організма в цілому, знаходяться в головному мозку. Пейсмекер часто називають біологічними часами, під якими розуміють властивість живих організмів орієнтуватись у часі. В ритмічній активності клітин домінуюча роль належить ядру. Ведучими циркадними ритмами у людини і тварин є ритми активності і спокою, сну і бадьорості і відповідні цим станам циклічні процеси витрачання і відновлення енергії. Одні вчені вважають, що циркадні ритми - це популяційно-клональна генетично закріплена періодичність, інші вважають, що ніякої генетичної закріпленості не існує.
Сезонні коливання сонячної активності, кількість вітамінів у їжі грають роль пускових механізмів у реалізації перечислених біоритмів. З цієї точки зору можна пояснити сезонність інфекційних хвороб, періодичність загострень виразкової хвороби шлунка і дванадцятипалої кишки, ревматизма, туберкульоза, не говорячи вже про такі соматичні хвороби, як гіпертонічна хвороба і інфаркт міокарда. Сезонні коливання проявляються у всіх процесах життєдіяльності. Підкреслюються, наприклад, ритми народження максимумів в весняний і мінімумів в осінньо-зимовий періоди і ритми смертності з зворотньою сезонною спрямованістю, останнє пов’язують з підвищенням чуттєвості до інфекцій в холодний період року.
Серед багаторічних циркаритмів краще інших вивчені цикли з 3х-річною періодичністю у хлопчиків і 2х-річною у дівчат, що проявляються з 10-літнього віку. “Психологічні ” ритми з періодом в 7 років описані Н.Я. Пєрном (1925). З окремою періодичністю протікають вікові розлади функцій, частково адаптації ока до яркого світла погіршується майже в 2 рази через кожні 13 років починаючи з 16 років.
Багаторічна періодичність явищ пов’язана з метеорологічними і геліогефізичними явищами (в тому числі флуктуаціями магнітного поля), які зумовлені дією сонцедіяльності (Чижевський, 1976). Сонячна активність зростає, коли її центри проходять через центральний меридіан поверненої до Землі напівсфери. Відомі 5-6, 11-12-річні і вікові цикли сонячної активності, але конкретний механізм біологічної дії космічних факторів не з’ясований. Припускається, що вони реалізуються через зміни фізико-хімічного стану коллоїдів (Піккарді, 1967). За гіпотезою С. Арреніуса, космічні впливи на земні явища сказуються через коливання електричного і магнітного полів Землі. Ряд вчених, проте, вважають ці коливання недостатніми для сприйняття організмом. Для того, щоб довести їх біологічну значимость, А.С. Пресман (1968) виказав ідею інформаційного впливу коливань електричних і магнітних полів. Ним допускається наявність в організмі внутрішніх систем електронно-магнітної регуляції чи вроджених осцилляцій (коливань), що мають різні періоди, амплітуду і сприймаючих коливання електричного і магнітних полів навколишнього середовища. Природньо, різноманітні зміни в організмі повинні бути циклічними, так як саме вони відбивають суть живого, забезпечують нормальну регуляцію життєвих процесів і рівновагу з навколишнім середовищем.
Але, як звісно, періодичні зміни властиві не лише людському організму, але і навколишньому середовищу. Ритми середовища існування називаються синхронізаторами - фізичними і соціальними.
Під фізичними синхронізаторами розуміється чергування світла і темноти, напруги електричних і магнітних полів, добові і сезонні коливання температури і вологості повітря, атмосферного тиску і т.д. Під соціальними - розпорядок виробничої і побутової діяльності.
І фізичні, і соціальні синхронізатори, постійно взаємодіють з біоритмами людського організму, накладають на них власний відбиток. Взяти хоча б біоритми функції нирок. За накопиченими даними, показники біоритмів ниркової функції залежать від умов праці і змінюються у людей, адаптованих до роботи в нічну зміну. А добовий ритм виділення води, іонів натрія, калія і хлора різний в різних географічних зонах.
1.3 Добові біологічні ритми
З усіх біоритмів людини найбільш вивчені і найбільш повно використовуються в практиці добові біоритми. Вони є основними в ієрархії часової організації людського організма. Стан системи добових ритмів організма є одним з основних критеріїв резистентності організма. Циркадний ритм фізіологічних процесів і поведінки спостерігається у всіх живих істот, який погоджений с добовою періодичністю змін метеорологічних факторів (освітленість, температура і атмосферний тиск). Поворот Землі навколо власної осі, зумовлює ритмічну зміну метеофакторів, визначає добовий ритм живої матерії. Цей ритм дуже стійкий. Значення динаміки циркадної ритміки фізіологічних показників є цінним діагностичним і прогностичним прийомом, особливо в процесі пристосування до різних клімато-географічних умов. Вивчення добової ритміки фізіологічних функцій організма є важливим в проведенні ефективної профілактики і лікуванні ряду хвороб. Вибір часу доби для проведення найбільш ефективного лікування має велике практичне значення. Циркадні ритми процесів організму синхронізовані з клімато-географічними, соціальними і іншими умовами навколишнього середовища. У випадку зміни умов навколишнього середовища відбувається розбіжність між астрономічним часом і фізіологічними ритмами, десинхроз, відновлення добової ритміки фізіологічних показників може бути критерієм видужання. Вивчення структури і фаз циркадної ритміки в процесі адаптації до постійно змінюючихся умов середовища дасть можливість знайти засоби для підвищення загальної резистентності організма до стресових ситуацій. Зовнішнє середовище динамічне, динамічні і функції організма.
Людина частіше за все і з’являється на світ і помирає вночі. Хворий, діждавшись ранку, загалом впевнений, що доживе до слідуючої ночі. Аварії і нещасні випадки на виробництві - їх теж більше вночі. Ще в 1729 році французький астроном де Меран довів, що рослини можуть відраховувати час - їх листки здійснюють окремі рухи протягом доби. Птахи, риби і комахи виробили у себе дивовижне відчуття часу. Вони можуть помилятися лише на декілька хвилин.
Нормальна структура добової ритміки показників вищої нервової діяльності і вегетативних функцій є однією з найважливіших параметрів адекватного функціонування організма до добових змін умов середовища і зумовлена прочністю механізмів адаптації. Відхилення від неї свідчить про порушення злагодженності і координації різних показників вищої нервової діяльності і вегетативних функцій, рівня регуляції функціональної діяльності і інтенсивності окисних процесів організма. Максимальний, середньодобовий і мінімальний рівень параметрів фізіологічних функцій означає відповідну взаємодію центрального і периферичного рівня регуляції систем.
Широко розповсюджена слідуюча точка зору, яку висловлювали Ашофф, (1964), Питтендрай (1964), Халберг (1964), ритм живої істоти за своєю суттю не є добовим. В постійних умовах, де не має нав’язаної зовнішньої періодичності, проявляється його власний період, величина якого залежить від рівня освітленості і температури. Якщо ж організм піддати чергуванню світла і темноти, вступає в дію механізм зсуву фази ритма, в результаті чого ритм затягується цією зовнішньою періодичністю і приймає нав’язаний їй період. При добовому циклі умов цей змушений період рівний 24 год. Отже, добовий ритм - це один з можливих випадків настройки ритма. Тому, всі ці ритми, включаючи добовий, називати циркадними. Але, не добовий ритм перетворюється в 24-годинний під впливом зовнішньої цикліки, а, навпаки, 24-годинний ритм під впливом неприродніх постійних умов світла і температури змінюється. Отже, циркадний ритм являє собою якби артефакт добового. На цю користь свідчить той факт, що в природі спостерігаються лише 24-годинна ритмічність. Циркадні ритми в природніх умовах майже неможливі. Є єдина доповідь про циркадну ритміку у деяких риб в умовах літнього полярного дня (Muller, 1973). Постійні умови - різкий і неприродній вплив на організм. В таких умовах істотно знижується життєможливість, наприклад, у деяких комах (Чернишев, Афонина, 1975), причому постійне світло і постійна темнота майже однаково згубні. Тому, важко погодитись з точкою зору, що постійні умови - деякий рівний нейтральний фон, на якому розгортається вільно протікаючий ритм. Також добовий ритм зберігається у деяких комах при так званому вільному виборі умов (Чернишев, Зотов, 1975). Якщо при примусовому постійному освітленні чи примусовій постійній темноті ритм у цих організмів циркадний, але в альтернативній камері, одна половина якої затемнена, а інша - постійно освітлена, тварини можуть вільно вибирати умови в даний проміжок часу умов освітлення, а ритм стає добовим.
Також доказом того, що саме добовий ритм лежить в основі виміру часу, слугують спостереження Ашоффа і Піттенгріфа та Дана, що демонструють стійкість ритму до зовнішніх впливів тим більше, чим ближче період ритма до 24 год.
На сьогоднішній час прийнято застосовувати термін “циркадний” для означення як істинних 24-годинних ритмів, так і ритмів, період яких відхиляється від 24 год. Таке об’єднання під однією назвою нормальних і перекручених добових ритмів, призводить до неправильної трактовки результатів. Тому, доцільніше буде називати терміном “добовий” лише 24 годинний ритм, а терміном “циркадний” - лише ритм, період якого відхиляється від 24 год під впливом постійних умов. Добові ритми можуть спостерігатися як в природі, так і в лабораторії, а циркадні - практично тільки в лабораторії.
Термін “циркадний” стали використовувати в широкому розумінні також тому, що в англійській мові відсутнє слово, рівне за змістом слов’янському - “добовий”. Але іноді застосовується термін “ diurnal" означає одночасно і добовий, і денний. Термін “ diel” - добовий, що запропонував Карпентер (1938), давно забутий і даремно.
Якщо добовий ритм являє собою тільки один з можливих варіантів циркадного ритму, то тоді в практиці можна використовувати різноманітні режими умов, період яких входить в інтервал периодів циркадного ритму. Проте, дослідження (Алякринський, 1977; Степанова, 1977; Стругхольд, 1977) доводять, що відхилення від 24-годинного режиму шкідливі для організму в цілому.
Добовий ритм, що спостерігається в природі складається з двох компонентів: ендогенного (добова організація) та екзогенна (різноманітний вплив на активність, як регулюючий ритм, так і безпосередньо впливаючих на поведінку).
В залежності від природи об’єкта питома вага цих компонентів різна. Найбільш виражений ендогенний ритм у таких тварин, активність яких пов’язана з необхідністю зміни середовища існування: вихода з укриття, з ґрунту, з води в повітряне середовище, з іла в водне середовище і т.д. Якщо ж він не змінює середовище існування, його ендогенний ритм виражений слабо.
В природніх умовах не виникає суттєвих протирічь між ендогенними ритмами і зовнішніми ритмічними впливами. Штучне порушення екзогенного компонента ритма призводить до порушення часової організації всієї живої системи і може визвати патологічні зміни функцій (Pittendrigh, Minis, 1972).
З дослідів Чернишева і Афоніна (1975) видно, що постійне світло чи постійна темнота і неприродній світловий режим пагубно сказується на тривалості життя мух дрозофіл і майже не впливає на жуків-короєдів. Цікаво, що ритм дрозофіл в цих умовах також порушується, в той час коли ритм жуків-короедів є більш стійким. Отже, ендогенний компонент ритма допомагає організму протидіяти неприроднім умовам, і чим більше вона виражена, тим стійкіше організм.
Ендогенний ритм дозволяє також рослинам і тваринам “передбачати” зміни умов і, таким чином, раніше підготовлятися до них. Крім того, ендогенний ритм перешкоджає реакції на несвоєчасну “провокаційну" зміну умов.
Ритм допомагає розмежуванню у часі активності тварин різних видів, призводячи до більш повного використання ресурсів. Різниця в ритмах у різних статей перешкоджає зхрещуванню між близькими родичами. Так, самці комах виходять з куколок, як правило, раніше за самок (Hardeland, 1969). Ще до вихода самок вони встигають відлетіти. З іншої сторони, синхронний виход зразу багатьох особин дозволяє безхребетним тваринам зустрітися при найменьшій втраті особин від метеорологічних умов і хижаків.
Ендогенний компонент ритма має велике значення в просторовій організації тварин. Властивість враховувати рух небесних тіл по небосводі для просторової організації була відкрита майже одночасно Фрішем у бджіл і Крамером у птахів. найбільш часто спостерігається орієнтація за сонцем. Біологічне значення такого астротаксіса очевидно. При астротаксісі відбувається як би перемикання тих ділянок сприймаючої поверхні ока, які повинні один за одним сприймати зображення небесного тіла. Хронометром тут є ендогенний ритм.
Крім просторової організації, бджіли використовують ендогенний ритм, пристосовуючись до розкладу виділення нектара різними квітами. Вони можуть запам’ятати час коли, отримують корм, і прилітають за ним кожного дня в окремі години доби. Такого роду поведінку, що спостерігається у багатьох тварин, називають рефлексом на час (Лобашев, Савватеєв, 1967).
Також, ендогенний добовий ритм має велике значення в сезонному фотоперіодизмі у рослин і тварин. Один з можливих способів визначення довжини фотоперіода заснований на порівнянні ендогенних ритмічних процесів з світловим циклом (Saunders, 1976).
Нормальна структура добової ритміки показників вищої нервової діяльності і вегетативних функцій є одним з найважливіших параметрів адекватного функціонування організма до добових змін умов середовища і зумовлена механізмами адаптації.
Важливим для людини є добовий ритм, тому що в цьому ритмі зосереджено майже 150 фізіологічних функцій людського організму. Ритмічний механізм з точки зору біологів заводиться регулярно зміною світла і темноти. Світло, попадаючи на сітчатку ока, через зорові нерви попадає в відділ головного мозку, під назвою гіпоталамус. Це вищий вегетативний центр, що здійснює складну інтеграцію і пристосування функцій внутрішніх органів і систем в цілісну діяльність організма. Він пов’язаний з гіпофізом, що регулює діяльність інших залоз внурішньої секреції, що виробляють гормони. Так, в результаті цього ланцюга кількість гормонів коливається в ритмі “світло - темнота”. Ці коливання і визначають високий рівень функцій організма вдень і низький вночі. У людини і тварин існують два механізми виміру часу: вроджений, заснований на системі внутрішнього годинника і взаємодії клітин, і механізм умовних рефлексів на час. Перший механізм здійснюється гіпоталамо-гіпофізарною системою, другий - корою головного мозку.
Доведено, що всі показники життєдіяльності організма (фізіологічні, біохімічні, етологічні) проявляються добовими коливаннями в онтогенезі. Різні ритми з’являються в різному віці. Народження дитини є поштовхом до виникнення добової ритміки фізіологічних процесів. Але, так як, людина живе не в ідеальних умовах, непостійними є фактори зовнішнього середовища, а також різноманітні фізичні і розумові навантаження, то і біоритми будуть постійно змінюватись, пристосовуючись до нових умов. Зміна добової ритміки фізіологічних показників залежить від рухової активності людини, від живлення (своєчасність і калорійність), від клімато-географічних умов середовища (атмосферний тиск повітря, ґрунту, швидкості вітра, вологості повітря і т.д.). ритм сну і бадьорості у новонароджених дітей швидко перетворюється в рефлекс на годування, але однофазності циркадного ритму, за своєю суттю є закріпленою філогенетично. Інші автори вважають, що ритмічна діяльність носить вроджений характер, вони виявили добові ритми вже на 1-й день життя навіть у недоношених дітей.
Отже, опираючись на літературні дані, можна зробити висновок, що добова ритміка фізіологічних систем організму проявляється не в один і той же час. Так, добова ритміка частоти пульса відмічається лише з 6-го тижня життя, температура тіла з 4-8-го тижня, ознаки добового ритму виведення з сечою натрія і калія відмічаються на 4-20-му тижні, а креатина і хлоридів - на 16-22 тижні після народження. Вірогідо, це залежить від ритму діяльності тієї чи іншої системи, і чим вище ритм, тим швидше утворюється рефлекс на час.
Добові ритми зміни більшості фізіологічних функцій у людини виражаються в постійному підвищенні їх рівня з ранкових годин до вечірніх (з максимальним підвищенням до 16-20 год) і подальшим зниженням в нічні години (з мінімальним значенням до 3-4 год. ранку). Деякі процеси (розпад гемоглобіна, коливання еозинофілів в крові, концентрація вуглекислого газу в крові, накопичення печінкою гемоглобіна і білка і т.д.) мають протилежну динаміку: їх максимальний рівень приходиться на нічні години незалежно від того, спить людина чи виконує якусь роботу.
Кожна температура досягає максимума до 18-19 год., а до 24-ої години знижується, мінімальне її значення приходиться на 3-4 ту год ночі. Різниця між денною і нічною температурою, її добовими коливаннями складає 0,8-1,3°С. Ректальна температура у дітей має два максимума: в 11-15 і 19-21 год. - і один мінімум - в 3-4 год. Порівняння температури різних ділянок тіла у дорослих показало, що якщо ректальна температура підвищується до 15 год. і потім знижується до 3 год., то температура на шкірі лоба змінюється трошки пізніше і не так виражено, а температура на шкірі верхівки середнього пальця змінюється протилежно першим двом. Отже, температура тіла і температура шкіри голови і тулуба повторюють добовий хід змін внутрішньої температура організма, а коливання температури шкіри кінцівок здвинуті на 180°, що пояснює зміни регуляції поверхневого кровообігу.
Добова періодика газообміну: вдень потреба в кисні максимальна, вночі - мінімальна. напруга СО2 крові, навпаки, максимальна в нічні години, до 3-4 год. вона може досягти 50 мм рт. ст. В добовому ритмі змінюються багато параметрів дихальної системи: частота дихання, сила вдоха, ЖЄЛ, збудливість дихального центру і т.д.
Функціональна активність серцево-судинної системи звичайно зростає з 8 до 12 год., а потім з 13 до 14-15 год. дещо знижується і знову зростає, досягаючи максимума до 18 год. Ввечері функціональна активність ССС знижується, її мінімум приходиться на 3-4 год. ночі. Частота пульса максимальнав 16 год., мінімальна - в 3 год. ночі. Артеріальний тиск в ранкові
25 години понижений, до 10-11 год. спостерігається його підйом, другий підйом приходиться на 16-20 год., після чого рівень артеріального тиску падає. Протягом доби закономірно змінюється об’єм циркулюючої крові: вдень зростає, а вночі знижується. Пульсове кровенаповнення мозку мінімально вночі, а максимально в 16-20 год., в тому ж напрямку коливаються показники венозного, мінімального, бічного і систолічного тиску крові в артеріях.
Робота серця також вдень протікає більш інтенсивно, чим вночі; систолічний і хвилинний об’єм крові максимальні ввечері (21-23год.) і мінімальні вночі. Відповідно змінюється довжина фаз систоли і ряду параметрів ЕКГ: довжина інтервалів PQ, QRS, QT і змінюється амплітуда зубців P, R, T. Добовий ритм ССС пов’язаний з тим, що вдень посилюється тонус симпатичної нервової системи, а вночі - парасимпатичної. Циркадні зміни рівня ССС зберігаються як при різкому зменшенні денної м’язової активності, так і при нічній праці.
Дослідження травної системи людини також виявили чітку добову періодику слиновиділення, шлункової і підшлункової секреції, моторики шлунка, секреторної функції печінки, зміни в печінці запасів глікогена, РНК і білка. Якщо вдень найбільш активні фізіологічні системи, що забезпечують його активну діяльність, то вночі - системи відновлення затрачених за день ресурсів. Так, накопичення в печінці глікогена, білка і РНК максимально в нічні години. У відповідності з біоритмами процесів травлення, протягом доби змінюється і склад в плазмі крові вільних амінокислот, вуглеводів, іонів К і Са, концентрація заліза. Описані внутрішньоклітинні коливання рівня нуклеїнових кислот і глікогена. Виражений добовий ритм частоти мітозів. В крові змінюється кількість еритроцитів, лейкоцитів і тромбоцитів. Змінюється також в’язкість крові, швидкість її зсідання, час склеювання тромбоцитів. В плазмі крові коливається склад хлора, фосфора, цинка, заліза,
26 натрія, калія і інших мінералів, змінюється рівень біологічно-активних речовин і гормонів. В роботі всієї ендокринної системи чітко виражений добові біоритми. Протягом доби змінюється концентрація в плазмі крові тестостерона, фолікулостимулюючого гормона, гормона росту, кортізола, серотоніна, тірозина і інших гормонів. При цьому час, що приходиться на максимум виділення гомонів ендокринними системами, не завжди співпадає: коливання їх рівня знаходиться в окремих фазових відношеннях. Так, гормони гіпофізарно-адреналінової ситеми мають найбільш високий рівень активності в ранкові години, а гормони росту - в нічні.
Коливання функціонального стану м’язової системи протягом доби визначають зміни рівня фізичних можливостей людини. Протягом дня м’язова сила зростає, протягом ночі - падає. Її максимум приходиться на 16-20 год., а мінімум - на нічні години. Витривалість до м’язової напруги (статична витривалість), м’язовий тонус, степінь тугості м’язів, внутрішньом’язовий тиск, електрична активність і інші показники характерізуються добовими показниками - підвищенням протягом дня і спадом протягом ночі.
Серед всіх циркадних біоритмів фізіологічних функцій організма найбільш виражена різниця денного і нічного рівнів активності, що властива центральній нервовій системі. Зміна функціонального стану нервової системи пов’язана з коливаннями збудливості і лабільності, швидкості проведення збудження і аккомодації. Збудливість кори головного мозку має протягом дня 2 підйома: 10-12 і 17 - 19год., які розділені меншим (денним) і великим (нічним) періодами її пониження. Протягом доби періодично змінюються частотні характеристики електроенцефалограми: енергія високочастотних спектрів ЕЕГ більш висока в ранкові години. Протягом доби також коливається рівень уваги, короткочасної пам’яті, швидкості підрахунку складів речення в друкованому тексті і т.д. В залежності від часу доби змінюється і стан сенсорної системи, в тому числі і порог світлової, кольорової і електричної чуттєвості ока, його здатність до аккомодації, критична частота мелькань, вірогідність знайдення звукового сигналу порогової інтенсивності, поріг електричної чуттєвості шкірних рецепторів і багато іншого. Так, вранці і вдень людина краще бачить, чує і розрізняє кольори, чим ввечері і вночі.
Фізіолог І.П. Павлов писав: “Не має нічого більш властного в житті людського суспільства, чим ритми”. Пристосуваність нервової системи людини до заданого ритму має велике значення в боротьбі з перевтомою, збереженні працездатності на довгі роки і адаптації до умов навколишнього середовища.
2. Поведінкові реакції тварин і методи їх вивчення
2.1 Форми поведінки тварин
Поведінка є одним з визначальних властивостей тваринного типу організмів. Вона розглядається як складна форма активності, для якої притаманні такі якості як рухи, спрямованість на підтримку сталості внутрішнього середовища, крім того, поведінка тварин реалізує нову вищу психічну форму відзеркалення.
Психіка тварин виникла із загальної властивості живих організмів - подразливості та зазнала значного біологічного прогресу, пройшовши шлях від сенсорної (відчуття окремих подразників) до перцептивної (синтез відчуттів та виникнення сприйняття та уявлень), потім до інтелекту (міркувальна діяльність тварин) та, нарешті, до свідомості людини, яка проявляється через мовну діяльність [22].
Структурно усі ці психічні форми відображення забезпечувались ускладненням будови нервової системи, яка пройшла шлях від дифузної до гангліонарної, а потім до центральної нервової труки з одночасним розвитком органів чуттів.
Таким чином, поведінка - це специфічний для тварин вид життєдіяльності, в якому досягається сталість внутрішнього середовища організму шляхом пошуку в зовнішньому середовищі біологічно значимих факторів, здійснення обміну інформацією, забезпечуючи адаптацію.
Гомеостаз та інформація - це дві головні функції поведінки. Кожний засіб пристосування тварин до умов середовища здійснюється шляхом певних функцій організму, забезпечуючи гомеостаз. Для цієї мети існують поведінкові реакції, які задовольняють потреби обміну речовин та інформаційні процеси в тваринному світі. В зв’язку з цим, репертуар
29 поведінкових реакцій включає локомоцію, їжедобування, терморегуляцію, підтримку чистоти тіла та виділення, пошук та організацію сховищ, пошукова активність, ігрова діяльність, репродуктивна активність [22].
Але поведінка базується не тільки на системах біологічної саморегуляції, також вона закодована в генетичному фоні виду, як звичайні процеси, які обумовлюють інші функції організму. Пластичність поведінки тварин є більш складною, бо вона генетично фіксована та передаються нащадкам не окремі готові рухи або їх поєднання, а норми реагування, в межах якої формуються рухові реакції в онтогенезі [24]. За даними Вагнера, який довів, що вродженими є не інстинктивні дії, а ті рамки, в межах яких ці дії можуть виконуватися в модифікованому вигляді відповідаючи певним умовам середовища. Реалізація вродженої програми поведінки в певних умовах індивідуального розвитка забезпечується процесами навчання. Виникає питання: “Що отримує тварина у “спадщину" від попередніх поколінь в генетично фіксованому вродженому вигляді? ” Чи має тварина самостійно оволодіти в порядку придбання індивідуального досвіда? К. Ліойд - Морган в своїй роботі “Привычка и инстинкт” (1899), дав аналіз вродженого та набутого в поведінці тварин, який побудований цілком на онтогенетичному підході. Він писав, що “діяльність, яка є результатом координування підібраних (в результата індивідуального досвіду) 10%первісно незв’язаних рухів, є новий продукт і цей продукт виступає результатом засвоєння, здобування, а не успадкування в якості певних, координованих рухів. Шляхом здобуття створюється “програма рухів" з даної кількості невизначених рухів, зкоординованих в певні відносини між собою. здобувається певна, координована, реактивна або відповідальна дія. Але, нараховуються відповідальні дії, які визначені з самого дня народження, які успадковуються готовими та сполучення, або координування яке відразу
30 після народження вже відрізняється повною досконалістю - в цьому випадку це не індивідуальні, а запозичені від предків”.
2.2 “Відкрите поле”, як один із методів вивчення оведінкових реакцій тварин
Методика “Видкритого поля”, яка широко використовується в сучасному нейрофізіологічному експерименті, отримала визнання завдяки своїй простоті та надійності.
Впроваджена в практику лабораторних досліджень Hall C. S. у 1934 році, вона дозволяє кількісно оцінити моторну активність щурів за 2-ма компонентами: вертикальній стійці та горизонтальній - локомоція. Крім того, дозволяє реєструвати окремі форми поведінки (захід у центр, нерухомість, кількість полюсів та інше) при всій важкості інтерпретації показників, що реєструються, може дати точні показники - характеристику стану тварин (страх, збудження або збільшення орієнтовної реакції) [15].
О.Г. Кенунен, В.Л. Козловський відмічають взаємозалежність між локомоцією та стійкою. Звичайне збільшення одного показника супроводжується збільшенням іншого. В нормі рухова поведінка щурів має певну “структуру”, що проявляється присутністю кореляції між кількістю стоєк та локомоціями.
Виявлений зв’язок між вертикальними та горизонтальними компонентами не залежить від рівня рухової активності. Співвідношення залишається при багаторазовому тестуванні [11].
Можна зробити висновок, що аналіз зв’язку між вертикальним та горизонтальним компонентами рухової поведінки щурів говорить про певну структуру цього виду природньої активності тварин. Аналогічний зв’язок був з’ясований О.Г. Кенуненом, В.Л. Козловським між компонентами локомоторної активності також і у щурів. Обчислювання коєфіцієнта кореляції між стійками та локомоціями вносить в широко відому методику “Відкритого поля” нову інформацію про цілісність структури рухової поведінки лабораторних тварин.
Що є “Відкрите поле” в загальних рисах, та що воно дійсно вимірює?
В сутності тест складається з вимірів поведінки тварин, яких розміщують в незнайомий відкритий простір, втеча з якого неможлива, так як його оточують прозорі стіни. Прояв цієї поведінки залежить від дії на тварину різних тест-факторів, таких як:
А) стимуляція (збудження), як результат здивування - переляку при перебуванні в звичайних умовах середовища;
Б) збудження тварин при переміщенні їх у “Відкрите поле”;
В) знаходження в оточуючому середовищі експеримента, який складається з “Відкритого поля” та його оточення;
Г) увесь попередній досвід при тест - ситуаціях.
В результаті цей останній фактор значить, що серед інших параметрів, в тесті вимірюється “звикання" та ” навчання” в реакції на тест ”Відкритого поля”. Кожний фактор відображає певний диференційований ефект у тварини з врахуванням різного генетичного та досвідного фону кожної тварини. Будь-який поведінковий експеримент показує відображення реакцій тварин на зовнішні фактори. Значимість кожного окремо виявленого типу поведінки буде похідним від взаємодії цих факторів.
Апаратура, її розміри, форма, колір, підрозділ на висоту стін, щільність підлоги, запах, характер розташування “стартового міста”, присутність або відсутність та природа додаткових стимулів - все це може варіювати. Також, саме умови оточуючого середовища - характер, інтенсивність та розташування джерел світла, звука та запахів, а також обозрівання та позиція дослідника - носить варіюючий характер.
Важкість стандартизації ускладнюється обмеженістю літературних повідомлень, які б наводили деталі корисних технологічних повідомлень варіантів в достатньому об’ємі. Щодо фізичної характеристики апаратури, то найбільш часто зустрічається форма “Відкритого поля” - кругла, рідко квадратна та прямокутна. Виникає питання, при яких умовах “Відкрите поле” перестає бути “Відкритим полем”? Остаточний варіант прямокутної форми звичайно визначається як “прямий" та поведінка, виміряна на ній може бути аналогічною поведінці, виміряній у “Відкритому полі” іншої форми. (e. g. W. Poley & I. P. Royce 1970, P. F. Zimbardo & K. C. Montgomery 1957).
Недостатньо вивчено вплив форми “Відкритого поля” на поведінку. Більш важливим є його розміри, вони змінюються при внутрішньовидових дослідженнях, причому в результаті зміни розміра поля суттєво змінюються деякі аспекти поведінки: рухомість є більш податливою впливу змін в апаратурі та оточуючому середовищі ніж дефекація. В той час, як дефекація у щурів більш незалежна від розмірів арени (Broadhurst P. L. 1957, 1958, 1950), то їх рухомість збільшується зі збільшенням площи поля (Broadhurst P. L. 1957, K. C. Montgomery 1951). Великий розмір поля створює непропорційно велике збільшення рухомості при умовах слабкого освітлення (Blizard D. A. 1971). На поведінку тварин може впливати не лише поле саме по собі, але і оточуючі об’єкти. Рівень освітленості як фактор оточуючого середовища був вивчений Candland D. K & Nagy Z. M. (1969), які встановили, що яскраве освітлення викликає зменшення рухової активності. Valle E. P. (1970), з’ясував, що амбуляція та рерінг зменшуються при яскравому освітленні. Вивчена кількісна кореляція рівня освітлення з іншими незалежними факторами: так, Hivesey P. Z. & Egger G.I. (1970) з’ясували, що амбуляція при освітленні 9,3 1х була менша чим при освітленні 0,005 1х, у щурів віком 18-24 днів.
Роботи H.I. Eysenck & Broadhurst P. L. 1964; Ivinskis A. (1965) присвячені оточуючому шуму. Але в багатьох випадках це охоплює тільки фоновий шум в лабораторії. Іноді додавався шум приблизно 80 ДБ. Дія шуму звичайно пригнічувала локомоцію (e. g. D. Bindra & Spinner N. 1958).
Hivesey P. Z. & Egger G.I. (1970) з’ясували, що “білий шум" 90 ДБ підвищує і амбуляцію і дефекацію. Дійсно, будь-який раптовий голосовий шум може значно пригнічити локомоцію та навіть викликати довготривалу зупинку рухомості у різних видів тварин (M. A. Hover 1970; Cummins R. A., Walsh R. N; Burtz-Olsen O. E.). Кількість залежних параметрів для оцінки поведінки у “Відкритому полі” постійно зростала протягом часу, починаючи з робіт Холла та Кальвіна (Calvin & Hall 1934; Hall & Ballechey 1932), та досягла 30:
І - Поведінка
А) (Рухи) Переміщення тіла (всього або більшої частини)
1. Тип рухів
відстань, які долають за одиницю часу
час амбуляції
частота підводжень на задні кінцівки
намагання уникати (стрибки)
латентний період
час без рухів
2. локомоція
“відвідування" певних відділів поля (центр або перефіричні кути)
Б) (Рухи) Переміщення частин тіла
маніпуляція об’єктами
принюхування
драпання
риття
оральний автоматизм
вмивання
вокалізація
візуальне обслідування
ІІ - Вегетативна нервова система
дефікація
урінація
ЧСС
ЧД
ІІІ - Надниркова активність
аскорбінова кислота наднирок
плазменні кортикостероїди
IV - Електроенцефалографія
гіпокамальний тета-ритм
електроміограмма
В основному це поведінкові ознаки, фізіологічні параметри, більшість з яких - варіанти рухової активності (амбуляція), завдяки легкості кількісних вимірів і явної надійності та вірогідності для інтерпретації. Вимірювання амбуляції звичайно робиться шляхом просторових підрахунків або вимірюванням пропорцій часу, який був використаний на мабуляцію. Виміри проводяться згідно:
1) характеристики їх чергових прагнень (цілі);
2) локальність;
3) латентність.
Амбуляція, яка була спрямована на втечу виміряна Ehrlich A. & Burns N. (1958). Місцезнаходження на периферії та в кутах (Morisson B.I. & Tratcher K. 1969), місцезнаходження біля стін (Vall F. D. 1970) - були використані як показники праці. Активність також може бути вивчена за її відсутності, за допомогою 2-х основних параметрів - латенція та фрізінг (“застивання, заморожування”). Латентний період вимірюється часом від початку експеримента до прояву певного типу поведінки. Звичайно, в якості латентного періода використовують час за який тварина залишає стартовий майданчик (Ivinskis A. 1968; W. Poley & I. P. Royce 1970). Теоретично може бути використана латентність будь - якого залежного параметра, але дослідники дають перевагу амбуляції.
Іншим обчислювальним параметром у “Відкритому полі” є відсутність активності - фрізінг. Він широко використовується як на індикатор стану стреса. Певно, це і є поведінкова реакція, яка на філогенетичній сходинці проявляється як сприйняття загрози (специфічна для кожного виду тварин). Факторами фрізінгу виступають раптові зміни, переміщення оточення (середові дислокації), фізичне обмеження рухомості та присутність хижаків. У “Відкритому полі” фрізінг - відповідь на подразники, які наведені вище і специфічні для кожного виду, іноді проявляються більше 1 години. Фрізінг може супроводжуватися тахіпноє та серцевою аритмією (M. A. Hofer, 1970). Другим варіантом довготривалої нерухомості виступає сон та уявний сон, який був досліджений Fox’ом & Spencer’ом І. М (1969). Тварини, у яких відмічали довготривалу нерухомість у “Відкритому полі”, швидко зупиняли рухи, посилювалася дефікація та урінація, вони навіть тремтіли при перенесенних в стартовий бокс. Уявний сон також відмічав Cummins R. A. у щурів, у яких виявлялося тремтіння, яке може бути результатом тахіпноє в стані нерухомості. M. A. Hofer (1970), Zynn R. (1966), відмічали уявний сон, як результат перезбудження, частота сну збільшувалась зі збільшенням часу досліду та часом між дослідами. Це скоріше звикання, яке вказує на те, що звикання до реакції збудження може супроводжуватися швидким станом дрімоти та сну. В цей період дрімоти може легко виникнути нереалізована орієнтовна реакція. Суб’єктивно, можливо диференціювати залякування від зовнішньої загрози та від псевдо-сплячки за характерними ознаками, такими як: широко відкриті очі, м’язова ригідність (скованість) проти постійного розвитку замкнутих очей та розслабленої пози, що супроводжують псевдосплячку. Ці дві форми поведінки наводять приклад реакції переляку та викликаної реакції збудження. Тестом для цих гіпотез може бути реакція реєстрації ЕЕГ тому, що орієнтовна реакція супроводжується гіпокамальним Q - ритмом, тоді як реакція переляку супроводжується десинхронізацією в гіпокампі. Отже, нерухомість у “Відкритому полі” може бути індикатором високої напруги (стресу).
Слідуючим критерієм активності є підводження - рерінг (Ivinskis A., 1968). В комплексі з амбуляцією він відображає стійкі індивідуальні ознаки “неспецифічний рівень збудженості”, який корелює з короткохвильовою активністю гіпокампу та з іншими поведінковими ознаками, також із варіантами інших поведінкових реакцій, в залежності від індивідуальних відмін в соматичних функціях, таких як, швидкість росту, температура тіла, тепловіддача, перевага певних видів їжі, ендокринні функції (Hat L. & Collova-Hemon 1968; Martinek Z. &Hat L. 1969). Сидіння виявляється при умовах, які зв’язані з низьким рівнем активності та негативно корелює з амбуляцією та рерінгом (Prescott R. 1970). Параметри, які не відносяться до рухів всього тіла, досліджені неповністю. Сюди відносяться: драпання, скрежет зубами, ЕМГ, серцеві ритми, ЕЕГ, які використовувалися для дослідження поведінки. Скрежет зубами було використано Hudhes’ом (1969) як можливий індикатор турботи (збентеженість), на тій основі, що воно може символізувати “поведінку заточення зубів з метою захисту”. Дихання та ЕМГ були використані Hofer’ом (1970) в дослідженні довготривалої амбуляції. Чищення шерсті (грумінг) і умивання, характеризують один і той же вид поведінки та визначають низьку вірогідність (Ivinskis A. 1968) та негативний зв’язок з показниками високоактивних станів (Prescott R. 1970).
Можливих параметрів вегетативної нервової системи багато, але на практиці вони звичайно лімітуються легко вимірювальними екскреторними та серцевими функціями. Характерним проявом пасивно-оборонної поведінки у щурів при переміщенні їх у нове незнайоме середовище, є вегетативна реакція у вигляді дефекації та урінації, а також зміненні рівня рухової активності. До форм пасивно - оборонної поведінки у тесті “Відкрите поле” наведені найменування: реакція дефікації і реакція урінації, що визначаються як “емоційність”, а рухову активність - як “амбуляцію”. Відображення пасивно-оборонної поведінки щурів через їх емоційність дослідив Hall (1934). У 1942 році Холл і Клайн порівняли 2-і лінії щурів за агресивністю. Вони з’ясували, що неемоційні щури були більш агресивні ніж емоційні. В зв’язку з їх дослідами можливо провести деяку аналогію між емоційністю і пасивно-оборонною поведінкою, як між неемоційністю та активно-оборонною поведінкою. Подальший розвиток методика “Відкритого поля” отримала у працях Broadhurstа (1957), який застосував не лише яскраве світло, але і гучний звук. Він виявив, що коефіцієнт надійності цього тесту дорівнював 0,82. таким чином, Broadhurst і Holl провели вдалий експеримент та вивели 2 лінії щурів: емоційну, за їх термінологією “реактивну” (MR) та неемоційну, або “нереактивну” (MNR). Ці дві лінії щурів вірогідно відрізняються за багатьма морфологічними та функціональними ознаками (Broadhurst & Eysenck, 1964), а також було відзначено, що дефікація і рухова активність (амбуляція) мають полігенну природу успадкування (Broadhurst P. L. 1959) [14].
Як прояв вегетативної реакції дефікація залишається домінуючим індексом емоційності, значення якої було підтверджено факторним аналізом (Whimdey & Denenberg, 1967). Крім дефікації враховується урінація за частотою прояву, присутністю або відсутністю, кількості або якості елімітуючої суміші з дефікацією (Tobach, 1966).
3. Методи та матеріали дослідження.
3.1 Методика вироблення штучного циркадного біоритму у самців щурів лінії Вістар
Дослідження проводилося на 60 самцях щурів лінії Вістар віком 7-7,5 місяців, масою 290-420 г, довжиною 330-410 мм, тривалість життя 2,5-3,5 роки, іноді до 5-6 років. Статева зрілість - 1,5-2 місяці.
За характером дії та інтенсивністю освітлення на початку експеримента було сформовано 3 групи:
К - контрольна (інтактна) - тварини знаходились при дії природного освітлення;
Д-Н - при дії штучного освітлення на протязі 12 годин на добу: з 6 години ранку до 18 години вечора);
Д - при дії цілодобового штучного освітлення.
Експеримент тривав 5 місяців. Кожна група утримувалася в окремих приміщеннях. Тварини знаходилися в стандартних клітках по 5 самців в кожній, при харчуванні ad libitum, вживання води було вільне.
Для створення штучного освітлення використовувалися лампи накалювання потужністю 100 Вт. Кожна клітка з тваринами мала свою власну лампу, яка була розміщена від неї на відстані 0,5 м. Рівень освітлення в приміщеннях зі штучним освітленням складав 300 лк. [8]
Саме така інтенсивність освітлення дає можливість ліквідувати розбіжність між циркадними ритмами у тварин. Якщо використовувати її нижчі або вищі показники, це призведе до суттєвого зрушення ритму виділення мелатоніну і, таким чином, до розбіжності ритмів організму в цілому. [10]
3.2 Методика проведення тесту “Відкрите поле”
Методика “відкритого поля” проводилась з використанням устрою, який являє собою квадрат розміром 1м х 1м, обмежений бортами висотою 40см. Вся площа поля розкреслена лініями на 25 квадратів зі стороною 20 см. [14] Над центром поля на відстані 0,5 м розташована лампа накалювання потужністю 100 Вт. Устрій знаходився у затемненому приміщенні. Перед розміщенням у “відкрите поле” щурів поодинці саджали у коробку і через 10 хв проводили тестування.
Спостереження за тваринами відбувалися на протязі 3х днів, в один і той же час доби за однакових умов освітлення і температури, за відсутністю сторонніх запахів і шуму. Час експозиції кожної тварини у “відкритому полі” становив 5 хв. Виборка тварин з кожної групи становила по 10 самців.
В індивідуальній поведінці тварин реєстрували їх горизонтальну рухову активність (амбуляції, локомоції); вертикальну активність (рерінг, підведення на задні лапки); латентний період (латенція); дефікацію (кількість болюсів - фекальних шариків); грумінг (кількість вмивань) та урінацію. [12,13]
4. Поведінкові реакції щурів які утримувались у різних вітлових режимах
4.1 Поведінкові реакції самців щурів контрольної групи
1) У самців даної групи був відсутній латентний період. Взагалі, латентність характеризується часом від початку досліду до появи певного типу поведінки і стартової позиції (Ivinskis A., 1968, Poley W. &Royce I. P., 1970). У нормі латентний період збільшується на 3 день (Furechtgott E. at al., 1961), злегка знижується на 2 день (Ader R., 1968). В даному випадку відсутність лацентації самців контрольної групи свідчить про їх високу активність і швидку адаптацію до умов навколишнього середовища.
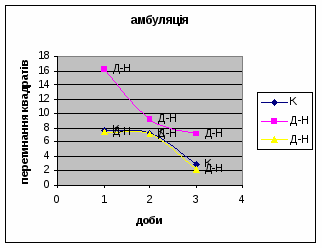
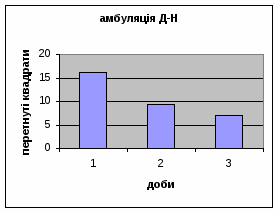
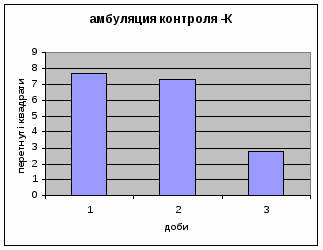
2) Амбуляція щурів цієї групи протягом експерементальних днів поступово зменшувалась з 7,68 до 2,82. (мал.1) (Амбуляція - локомоція - це кількість перетнутих квадратів твариною всіма чотирма лапами; це факторний комплексний параметр, що відображає дослідницьку діяльність і емоційність). Це говорить про наявність у них редукції страху. Отримані дані відповідають літературним. Адже, за Whimbey A. E., Denenberg’ом (1969) частота амбуляції через інтервал часу змінюється від позитивної - в 1й день, до негативної на 2й день і т.д. малюнок 1
3) Вегетативні показники.
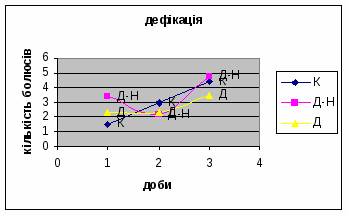
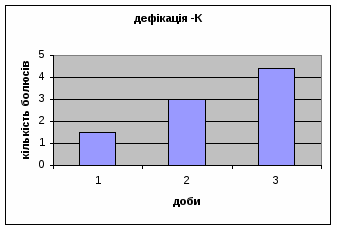
Дефікація зростала з 1,5 до 4,4 (мал.2), що відповідає літературним даним. Так, за Broadhurst’ом Р.Н. (1969) виявлено тенденцію до її зменшення протягом 3х днів спостереження. Загалом, дефікація та урінація - це перші показники емоційності тварин. Таким чином, отримані дані свідчать про те, що у самців цієї групи знижується емоційний стан і спостерігається розвиток звикання до умов. Відсутність латентного періоду та позитивна кореляція між амбуляцією та дефікацією, також свідчить на цю користь.
Малюнок 2
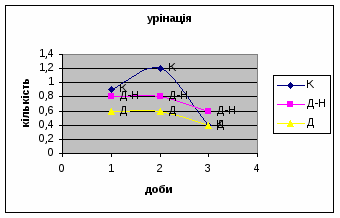
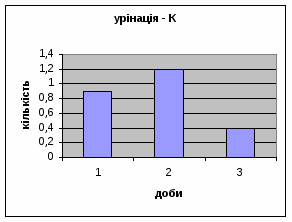
Спостерігалось зменшення показників урінації у самцув контрольної групи від 0,9 до 0,4 (мал.3), хоча на другий день експеременту спостерігалось деяке підвищення сечовипускання що. Отримані дані а також позитивна кореляція власне цього показника з амбуляцією, підтверджує думку про редукцію страху у цих щурів.
Малюнок 3
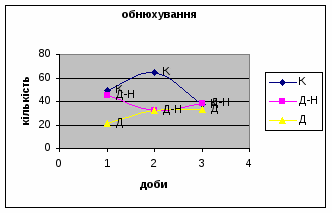
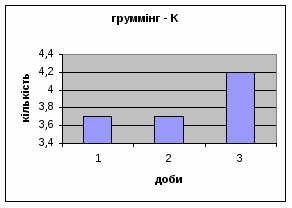
4) Протягом експерименту у піддослідних тварин зменшувались показники рерінгу (вставання на задні лапки, це основний показник активності тварин і використовується він як індекс активності) принюхування (мал.4 і мал.5), та зростали показники грумінгу (вмивання) (мал.6), що разом з описаними вище даними говорить про те, що у самців даної групи знижується емоційність і розвивається звикання до стресових факторів навколишнього середовища.
Малюнок 4 Малюнок 5
Малюнок 6
4.2 Поведінкові реакції самців щурів Д-Н групи
1) Відсутня лацентація.
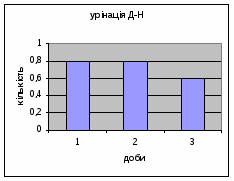
43 2) Амбуляція у самців Д-Н-групи протягом 3х днів зменьшувалась з 16,2 до 7,11. (мал.7) Тобто спостерігається більш поступове зниження цих показників в порівнянні з контрольною групою, і, в той же час, більша рухова активність, що проявлялась в перетнутих квадратах.
Малюнок 7
3) Вегетативні показники.
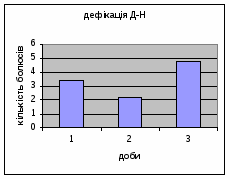
Показники дефікації, в порівнянні з іншими групами, мали свою криву: на другий день експерименту кількість болюсів різко зменшилась з 3,4 до 2,2 (мал.8), а потім почала поступово зростати до 4,8. Ці дані свідчать про те, що у самців Д-Н-групи відбувається більш повільніша редукція страху і емоційності, в порівнянні з іншими групами піддослідних тварин. Позитивна кореляція між амбуляцією і дефікацією також дає підстави вважати те, що існує зниження тривоги у даних самців і загальна їх адаптація до умов. Урінація протягом перших 2х днів експерименту була незмінна - 0.8, а далі почала знижуватись до 0,6 (мал.9).
Малюнок 8 Малюнок 9
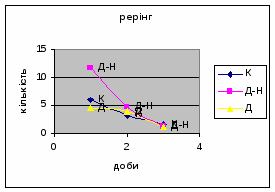
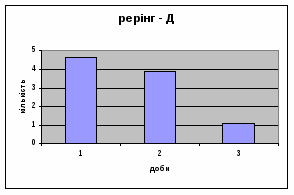
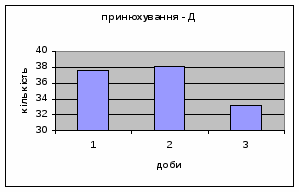
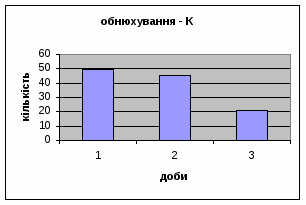
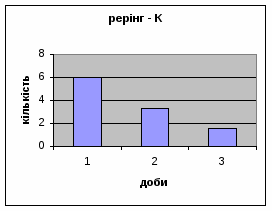
4) Також специфічність самців Д-Н-групи проявилась і при аналізі показників принюхуванні та рерінгу. Загальне число принюхувань, в порівнянні з іншими групами набагато вище, так в перший день дослідження зареєстровано 64,4 принюхувань. Як і в контрольній групі, даний показник зменшувався (до 32,3) (мал.11). Найбільший коефіцієнт активності (рерінг) був зареєстрований саме в цій групі тварин.
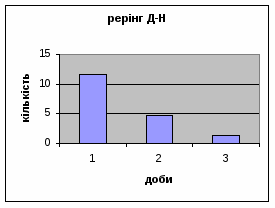
Протягом перших двох днів досліду він майже був незмінний - 11,7, а потім різко почав знижуватись аж до 1,3 (мал.10). Все вище описане говорить про те, що дані самці Д-Н-групи більш важче пристосовуються до умов середовища, особливо в перші дні, де адаптація майже не спостерігається. Але потім, ці піддослідні тварини активно наздоганяють в цьому аспекті тварин інших груп. Про що свідчить позитивна кореляція дефікації, амбуляції та урінації між собою, а також зменшення показників рерінгу та принюхування. Таким чином, у самців Д-Н-групи також відбувається редукція емоційності, тривоги, тобто розвивається звикання до стресових факторів.
Малюнок 11 Малюнок 10
4.3 Поведінкові реакції самців щурів Д-групи
1) Відсутня лацентація.
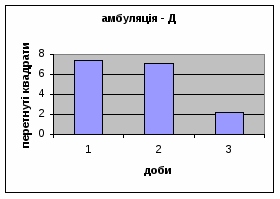
2) У самців Д-групи спостерігалась найнижча локомоція в порівнянні з тваринами інших груп. Це говорить про те, що світло для них був звичним фактором існування. На відміну від інших груп, серед переміщення даних тварин по “полю" було зареєстровано перетинання майже всіма піддослідними центра “поля”. Такий характер амбуляції не спостерігався серед інших експериментальних груп. Деякі Д-самці навіть тривалий час знаходились безпосередньо під лампою. Таким чином, на основі отриманих фактів, можна сказати, що тварини цієї групи майже не відчували страху бо дія світла для них була звичною. Показники амбуляції протягом 3х днів знижувалась з 7,38 до 2,19 (мал.12).
Малюнок 12
3) Вегетативні показники.
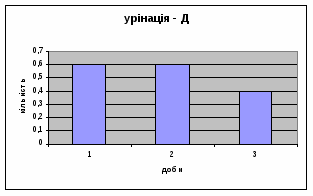
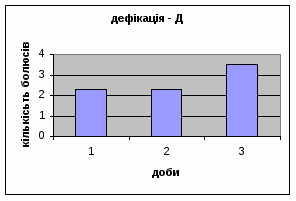
Показники дефікації зростали з 2,3 до 3,5 (мал.13), що говорить про швидке звикання до умов існування. Позитивна кореляція між показниками дефікації та локомоції свідчать також на цю користь. Найменша кількість урінацій (від0,6 до 0,4) (мал.14) була зареєстрована саме в цій групі самців, яка також мала позитивну кореляцію з амбуляцією.
Малюнок 12 Малюнок 13
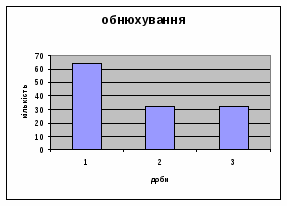
4) Самці Д-групи за показниками рерінгу були найменш активними серед експериментальних груп. Як і в попередніх групах дані показники цього параметру зменшувались (з 4,6 до 1,1) (мал.15). Але в порівнянні з К і Д-Н - групами, самці Д-групи протягом перших 3х днів експерименту проявляли стійки в напрямку до лампи, що не спостерігалось серед інших груп. Знову, це підтверджує думку про те, що дані тварини не відчували страху перед дією світла. Це, а також незначне зменшення кількості принюхувань (від 37,5 до 33,2) (мал14), дає можливість зробити висновок про швидкий розвиток у самців Д-групи звикання до стресових факторів середовища.
Малюнок 14 ёМалюнок 15
Висновки
1. У всіх групах експериментальних тварин було зареєстровано зниження амбуляції (мал.16) і підвищення дефікації (мал.17).
Малюнок 16
Малюнок 17
Але у самців Д-Н-групи відбувається більш повільніша редукція страху і емоційності, в порівнянні з іншими групами (К і Д) піддослідних тварин, так як, на другий день експерименту кількість болюсів у них різко зменшилась. У самців Д-групи спостерігалась найнижча локомоція, бо світло для них було звичним фактором існування. Також серед тварин цієї групи було зареєстровано перетинання майже всіма піддослідними центра “поля”, що не спостерігалось серед інших груп. Деякі Д-самці навіть тривалий час знаходились безпосередньо під лампою. Отже, тварини цієї групи майже не відчували страху, бо дія світла для них була звичною.
2. Загальне число принюхувань найбільше серед тварин Д-Н групи. На відміну від самців К - і Д - груп на другий день експерименту вони проявили найменшу нюхову активність - свій мінімум (мал.18), в той час як у останніх спостерігався її підйом, а далі поступове зниження.
Малюнок 18
3. Найбільший коефіцієнт активності (рерінг) був зареєстрований саме в Д-Н групі тварин (мал. 19). Отже, що дані самці більш важче пристосовуються до умов середовища, особливо в перші дні, де адаптація у них майже не спостерігається. Самці Д-групи за показниками рерінгу були найменш активними серед експериментальних груп. Але в порівнянні з К і Д-Н - групами, вони протягом перших 3х днів експерименту проявляли стійки в напрямку до лампи. Тобто, дані тварини не відчували страху перед дією світла.
Малюнок 19
4. Найменша кількість урінацій (від 0,6 до 0,4) (мал. 20) була зареєстрована саме в Д - групі самців. Загалом, у всіх групах вона поступово зменшується, що говорить про редукцію страху тварин.
Малюнок 20
5. Вище описані дані дають можливість стверджувати, що даним піддослідним самцям всіх груп властива емоційна, неагресивна, пасивно - оборонна поведінка (Кулагин Д.А., Федоров В.К., 1969). Тобто домінантними у поведінці даних тварин є стан психічної “відмови від пошуку”,“визнання поразки”. Як правило, наявність неспецифічних умов існування супроводжується появою психосоматичних захворювань. А це в свою чергу, при вище описаній формі поведінки може призвести до елімінації особин, перевага яких могла б негативно відбитись на життєдіяльності популяції вцілому, що сприяє еволюції виду і попереджає його регрес.
Отже, найповільніше адаптуються до стресових умов середовища самці Д-Н-групи, а найшвидше - самці Д-групи, так як вони підлягали цілодобової дії абіогенного фактора (світла) протягом 5 місяців, і за цей час встигли до нього звикнути. Про високу спроможність активно адаптуватись до стресових умов середовища свідчить і той факт, що у представників всіх груп був відсутній латентний період.
Література
1. Агаджанян Н.А. Биол. ритмы. - М.: Медицина., 1967. - 120с.
2. Ашофф Ю.А. Биол. ритмы: в 2х томах. Под ред. Ю.А. Ашоффа. - М.: Мир, Т.1. - 412с.
3. Биолог. ритмы. М.: Наука, 1980, 319с. Авт.: Ю.А. Романов,
С.А. Чепурнов, Г.А. Клевезоль.
4. Биологические часы. [Сб-к статей] Пер-д с англ. под ред. С.Э. Шноля. - М.: Мир, 1964. - 694с
5. Деряпа Н.Р. Проблемы мед. биоритмологии. - М.: Медицина., 1985. - 208с.
6. Дильман В.М. Большие биологические часы: Введение в интегральную медецину. - М.: Знание, 1981. - 208 с
7. Доскин В.А., Лаврентьева Н.А. Ритмы жизни. - 2-е изд. перераб. и доп. М.: Медицина, 1991. - 172с
8. Ефимов М.Л. Биол. ритмы в норме и патологии. - Алма-Ата. Казахстан, 1981. - 152с.
9. Зидерманс А.А. Некоторые вопросы хронобиологии и хрономедицыны: Обзор лит. / АН. Латв. ССР, Ин-т орган синтеза. Рига: Зинотне, 1988. - 212с
10. Лобашев М.Е. и Саватеев В.Б. Физиологические суточные ритмы животных.М. - Л. Изд-во Академии наук СССР, 1959. - 259с
11. Кенунен О.Г., Козловский В.Л. Структура двигательного поведения лабораторных животных - новые возможности методики “ Открытого поля”. // Физиолог. журн. им. Сеченова 1992 - Т.78, № 1
12. Комаров Ф.И., Рапопорт С.И. Хронобиология и хрономедецина. - М., “Триада-Х”, 2000. - 288с
13. Кузюта Э.И., Серохвостов А.П. Ритмы под солнечным ветром. - Фрунзе, Кыргыстан, 1991. - 104с.
14. Кулагин Д.А., Федоров В.К. Генетика поведения. - Ин-т физиологии им.А. Павлова. АН СССР. “ Наука”. 1969.
15. Маркель А.Л., Хусаинов Р.А. Метод комплексной регистрации поведенческих и вегетативных реакций у крыс при проведении теста “Открытого поля”. // Высшая нервная д-ть. 1976 - Т.26, №6.
16. Матюхин В.А. Биоритмология перемещений человека. - Новосибирск: “Наука”, Сиб. отделение, 1976. - 104 с
17. Морфофизиологический анализ формирования механизма цикла активность - покой в онтогенезе человека / А.И. Белич, М. Н. Константинова // Вестник Российской Академии мед. наук. - 1996 №3 ст55-61
18. Павлович Н.В. Биомагнитные ритмы. - Минск: Университетское, 1991. - 134с.
19. Путилов А.А. “Совы”, “Жаворонки” и др.: О наших внутренних часах и их влиянии на здоровье и характер. - Новосибирск: Изд-во Новосиб. ун-та.; М.: Совершенство, 1997. - 263с.
20. Ритмы жизни / Л.В. Львова. Провизор. - 2003 №1. - с.34-37
21. Руттенбург С.О. Циркадный ритм Физиологических процессов и трудовая деятельность человека. - Фрунзе: “Илим”, 1976. - 188с
22. Силантьева К.Г. Поведение и его механизмы: Учеб. пособие к спецкурсу [Поведение животных] / Урал. гос. пед. ин-т. - Екатеринбург: Урал. ГПИ, 1992. - 133
23. Уинфри А.Т. Время по биологическим часам / Пер. с анг. А.М. Алпатова; под ред.В.И. Кринского. - М.: Мир, 1990. - 208с
24. Фарби К.Э. Основы психозоологии: [Учеб. Длявузов по спец. “Психология”, “Биология”, “Зоология" и “Физиология”]. - 3-е изд. М.: Учеб. - метод. коллектор “Психология”, 2001. - 463.
25. Фельдман Г.Л. Биоритмология. Изд-во Ростовского ун-та, 1982. - 80с.
26. Хаснулин В.И. Космические тайны вашего самочувствия. Новосибирск: Наука. Сиб. отд-ние, 1992. - 176с.
27. Хронобиология и хрономедецина: Руков-во / Ф.И. Лимарова и др. под ред.Ф.И. Комарова. - М.: Медицина, 1989. - 399с
28. Hall C. S. 1934a Comp. Psychol., 17, p89.
29. Hall C. S. 1934b Comp. Psychol., 18, p385.
30. Hall C. S. and L. L. Klein. 1942.comp. Psychol., 33, p371.

Нравится материал? Поддержи автора!
Ещё документы из категории биология:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ