Процесс фотосинтеза

 Введение
Введение
Фотосинтез и первичная биологическая продуктивность
Физиологическая роль азота, круговорот азота в атмосфере
Поглощение минеральных веществ корнями растений
Дерево испарило за 1 ч 500г H2О, а корневая система поглотила за это время 450г воды. Какие условия внешней среды могли вызвать указанное несовпадение количества поглощенной и испаренной воды. Как это отразится на испарении
5. Библиографический список
1. Фотосинтез и первичная биологическая продуктивность
Фотосинтез (от греч. φωτο- — свет и σύνθεσις — синтез, совмещение, помещение вместе) — процесс образования органических веществ из углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилл у растений, бактериохлорофилл и бактериородопсин у бактерий). В современной физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества.
Фотосинтез является основным источником биологической энергии, фотосинтезирующие автотрофы используют её для синтеза органических веществ из неорганических, гетеротрофы существуют за счёт энергии, запасённой автотрофами в виде химических связей, высвобождая её в процессах дыхания и брожения. Энергия получаемая человечеством при сжигании ископаемого топлива (уголь, нефть, природный газ, торф) также является запасённой в процессе фотосинтеза.
Фотосинтез является главным входом неорганического углерода в биологический цикл. Весь свободный кислород атмосферы — биогенного происхождения и является побочным продуктом фотосинтеза. Формирование окислительной атмосферы (кислородная катастрофа) полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования озонового слоя, позволило жизни выйти на сушу.
Биологическая продуктивность, образование биомассы организмами, выражаемое потоками органического вещества и его потенциальной химической энергии на единицу площади за единицу времени. Понятие биологическая продуктивность применимо к растительности, сообществам (фитоценозам), к их отдельным ярусам, к отдельным популяциям растений и животных. Продуктивность всех популяций организмов на единицу площади характеризует биологическая продуктивность биогеоценозов и экосистем. Биологическая продуктивность количественно оценивают по её результату главным образом по годичной биологической продукции сухого органического вещества (в т/га • год, г/м2 • год), или энергии (Дж/м2 • год, ккал/га • год, кал/м2 • год). Специфику процесса изучает физиология растений. Биологическая продуктивность иногда отождествляют с запасами биомассы на единицу площади, что может лишь отчасти характеризовать биологическая продуктивность. Различают первичную и вторичную биологическую продуктивность. Первичная биологическая продуктивность характеризуется образованием биомассы (первичной продукции) в процессе фотосинтеза зелёными растениями (автотрофами), которые образуют первый трофический уровень экосистемы и служат началом всех цепей питания. К первичным продуцентам относят и некоторые хемосинтезирующие бактерии. В процессе утилизации вещества и энергии первичной продукции образуется биомасса всех гетеротрофных организмов (бактерий, грибов и животных), называемых консументами. Продукция консументов характеризует вторичную биологическая продуктивность, к которой относят и массу хищных животных, питающихся растительноядными и другими хищниками.
Первичная биологическая продуктивность.
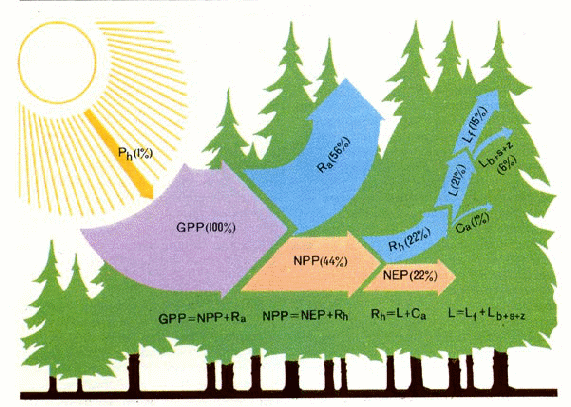
При исследованиях первичной биологической продуктивности наземных биогеоценозов определяют ряд показателей, которые затем используют в качестве отдельных статей баланса органические вещества на конкретных участках. Продукция, определяемая с учётом затрат вещества и энергии на процессы метаболизма самих организмов-продуцентов, называется первичной брутто-продукцией, или валовой продукцией (обычно обозначают GPP — от англ, gross primary production). Разность между первичной брутто-продукцией и затратами растений на дыхание (Ra) определяет первичную нетто-продукцию — NPP (от англ, net primary production). В лесном фитоценозе NPP включает в себя не только чистую продукцию прироста за учитываемый период (истинный прирост фитомассы) — NEP (net ecosystem production), но и продукцию, перешедшую за то же время в опад (листья, цветки, семена и др.) и отпад (отмершие деревья, сучья и др.), которые суммарно обозначаются L, а также часть продукции живых растений, пошедшую на корм животных-фитофагов (консумпцию) — Сa. Сумму этих показателей часто называют гетеротрофным дыханием (Rh), поскольку энергия в обоих этих потоках (Rh=L+Ca) освобождается главным образом с участием гетеротрофных организмов. Для консументов, независимо от их трофической специализации, применяют иную схему. Отчуждаемая при консумпции фитофагами продукция растений в некотором количестве поедается животными, остальная (огрызки, объедки) поступает в опад. Съеденная пища частично ассимилируется организмами, частично экскре-тируется и поступает в детрит. За счёт продуктов ассимиляции происходит прирост биомассы, т. е. формируется продукция и поддерживаются процессы метаболизма. В продукцию включаются вещество или энергия прироста (привеса) животных за изучаемый период и прироста потомства. Эти величины, с учётом вещества и энергии элиминированных особей, характеризуют прирост биомассы животных. Биомасса животных-иммигрантов в продукцию не включается. При этом ассимилированная пища и прирост биомассы животных соответствуют общей (брутто) и чистой (нетто) продукции автотрофов.
Первичная биологическая продуктивность зависит от интенсивности фотосинтеза растениями и продолжительности его периода, фотосинтезирующей поверхности фитоценозов и древостоев, выражаемой индексом листовой поверхности и характером расположения листвы в толще полога, а также от кол-ва поступающей фотосинтетической радиации, условий увлажнения и минерального питания. Кульминация первичной продукции насаждений, выражаемая чистой продукцией (NPP) или истинным приростом фитомассы (NEP), приходится на возраст 20—40 лет. Однако в искусств, насаждениях она наступает раньше, чем в естественных, хотя с возрастом различия Б. п. лесных культур и естественных древостоев сглаживаются.
2. Физиологическая роль азота, круговорот азота в атмосфере
Азот – биоэлемент, структурная единица органических соединений, участвует в построении организмов и обеспечении их жизнедеятельности. Входит в состав важнейших биополимеров: белков, нуклеиновых кислот (ДНК, РНК); некоторых витаминов и гормонов. В воздухе азота содержится 78% по объему и 75,5% по массе.
Азотфиксирующие бактерии способны усваивать азот непосредственно из воздуха, превращая его в аммиак. Они живут самостоятельно, например азотобактер, цианобактерии, азоспириллы, или поселяются в корнях бобовых растений (клевер, горох, люпин, вика и др.) – бактерии рода ризобиум. Над 1 га почвы в атмосфере содержится более 70 тыс. т свободного азота, и только в результате азотфиксации часть этого азота становится доступной для питания высших растений (содержание доступного для растения азота в почве очень невелико). При связывании N2 клубеньковыми бактериями в симбиозе с растениями семейства бобовых почва ежегодно обогащается азотом на 200–300 кг/га, а свободноживущие бактерии вносят в почву азота 1–3 кг/га в год. На рисовых полях свободноживущие цианобактерии фиксируют 30–50 кг молекулярного азота на 1 га в год. Известно довольно много азотфиксаторов: бактерии, актиномицеты, дрожжевые и плесневые грибы, синезеленые водоросли.
Растения поглощают азот из почвы в виде растворимых нитратов и солей аммония (NH4+). Соли транспортируются в стебли и листья, где в процессе биосинтеза очень быстро превращаются в аминокислоты и белки – неотъемлемую часть любого живого организма.
Азот составляет 0,3–4,5% от массы растения. Он усиливает рост стеблей и листьев. При недостатке азота замедляется рост растения, образование хлорофилла, листья приобретают бледно-зеленую окраску и преждевременно желтеют, стебли становятся тонкими и слабо ветвятся, вновь образующиеся листья мельчают, цветки, не раскрываясь, засыхают и опадают. При длительном азотном голодании бледно-зеленые листья приобретают желтый, оранжевый или красный оттенки.
Существуют растения-индикаторы, которые великолепно растут при повышенном содержании азота в почве. Это хорошо знакомые нам крапива, малина, чистотел, пырей ползучий.
Азот — незаменимый биогенный элемент, так как он входит в состав белков и нуклеиновых кислот. Круговорот азота один из самых сложных, поскольку включает как газовую, так и минеральную фазу, и одновременно самых идеальных круговоротов.
Круговорот азота тесно связан с круговоротом углерода. Как правило, азот следует за углеродом, вместе с которым он участвует в образовании всех протеиновых веществ.
Атмосферный воздух, содержащий 78% азота, является неисчерпаемым резервуаром. Однако основная часть живых организмов не может непосредственно использовать этот азот. Он должен быть предварительно связан в виде химических соединений. Например, для усвоения азота растениями необходимо, чтобы он входил в состав ионов аммония (NH4+) или нитрата (NO3-).
Газообразный азот непрерывно поступает в атмосферу в результате работы денитрофицирующих бактерий, а бактерии-фиксаторы вместе с сине-зелеными водорослями (цианофитами) постоянно поглощают его, преобразуя в нитраты.
Важную роль в превращении газообразного азота в аммонийную форму в ходе так называемой азотофиксации играют бактерии из рода Rhizobium, живущие в клубеньках на корнях бобовых растений. Растения обеспечивают бактерий местообитанием и пищей (сахара), получая взамен от них доступную форму азота. По пищевым цепям органический (входящий в состав органических молекул) азот передается от бобовых другим организмам экосистемы. В процессе клеточного дыхания белки и другие содержащие азот органические соединения расщепляются, азот выделяется в среду большей частью в аммонийной форме (NH4+). Некоторые бактерии способны переводить ее и в нитратную (NO3-) форму. Отметим, что обе эти формы азота усваиваются любыми растениями. Азот, таким образом, совершает круговорот как минеральный биоген. Однако такая минерализация обратима, так как почвенные бактерии постоянно превращают нитраты снова в газообразный азот.
В водной среде также существуют различные виды нитрофицирующих бактерий, но главная роль в фиксации атмосферного азота здесь принадлежит многочисленным видам способных к фотосинтезу сине-зеленых водорослей из родов Anabaena, Nostoc, Frichodesmium и др.
Круговорот азота четко прослеживается и на уровне деструкторов. Протеины и другие формы органического азота, содержащиеся в растениях и животных после их гибели, подвергаются воздействию гетеротрофных бактерий, актиномицетов, грибов (биоредуцирующих микроорганизмов), которые вырабатывают необходимую им энергию восстановлением этого органического азота, преобразуя его таким образом в аммиак.
В почвах происходит процесс нитрификации, состоящий из цепи реакций, где при участии микроорганизмов осуществляется окисление иона аммония (МН4+) до нитрита (NO2-) или нитрита до нитрата (NО3-). Восстановление нитритов и нитратов до газообразных соединений молекулярного азота (N2) или окиси азота (N2O) составляет сущность процесса денитрификации.
Образование нитратов неорганическим путем в небольших количествах постоянно происходит и в атмосфере: путем связывания атмосферного азота с кислородом в процессе электрических разрядов во время гроз, а затем выпадением с дождями на поверхность почвы.
Еще одним источником атмосферного азота являются вулканы, компенсирующие потери азота, выключенного из круговорота при седиментации или осаждении его на дно океанов.
В целом же среднее поступление нитратного азота абиотического происхождения при осаждении из атмосферы в почву не превышает 10 кг (год/га), свободные бактерии дают 25 кг (год/га), в то время как симбиоз Rhizobium с бобовыми растениями в среднем продуцирует 200 кг (год/га). Преобладающая часть связанного азота перерабатывается денитрифицирующими бактериями в N и вновь возвращается в атмосферу. Лишь около 10% аммонифицированного и нитрифицированного азота поглощается из почвы высшими растениями и оказывается в распоряжении многоклеточных представителей биоценозов.
3. Поглощение минеральных веществ корнями растений
Корень зеленого растения имеет свое характерное строение. Верхняя, наиболее старая, его часть покрыта слоем пробковой ткани и не участвует в питании растений, нижняя, молодая, - оболочками, легко проницаемыми для воды и солей. Растущий корень имеет 4 зоны.
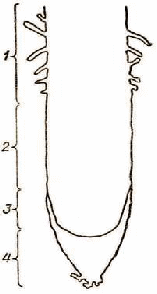
Самой молодой растущей частью корня является его кончик, покрытый снаружи защитным корневым чехликом (рис. 7). Корневой чехлик предохраняет нежные делящиеся клетки корневой меристемы от разрушения, способствует росту корня и проникновению его в глубь почвы. Клетки корневого чехлика содержат крахмальные зерна, которые редко используются растением для питания. Наружные клетки чехлика систематически стираются при росте корня и гибнут, образуя большое количество слизи, которая облегчает продвижение корня в почве. Внутри чехлика все время нарастают новые клетки, поэтому объем его не изменяется.
Рис. Продольный разрез кончика корня пшеницы:
1 - зона всасывания с корневыми волосками; 2 - зона роста; 3 - зона делящихся клеток; 4 - корневой чехлик
Непосредственно под корневым чехликом находится конус нарастания корня. Эта часть корня называется зоной делящихся клеток и по длине занимает всего несколько миллиметров. Выше зоны делящихся клеток находится зона роста, которая также составляет несколько миллиметров.
Над зоной роста (на участке длиной от 5 до 20 мм) расположена зона всасывания (или поглощения). Поверхность корня в этой зоне густо покрыта корневыми волосками - выростами клеток. Они служат для всасывания из почвы воды и минеральных солей; размеры их малы, и они различимы только с помощью оптического увеличения. Оболочки волосков пронизаны обычно тяжами протоплазмы, благодаря чему осуществляется непосредственный контакт внутреннего содержимого клеток с почвой. Корень непрерывно растет и образует все новые волоски, старые же быстро отмирают. Интересно отметить, что каждый корневой волосок живет не более 15-20 сут. Корневых волосков вырастает очень много, например на 1 мм2 поверхности корня кукурузы до 425, гороха - 230. Подсчитано также, что общая площадь поверхности всех корней вместе с корневыми волосками одного хорошо развитого куста озимой ржи может достигать 237 м2, что в 130 раз превосходит площадь надземной части этого растения. Общая длина всех корней одного отдельно растущего растения ржи составляет в среднем около 80 км, причем самые длинные корни уходят в почву на глубину до 1,6 м. При выкопке растения мы не обнаруживаем такого количества корней, так как при отделении земли значительная часть тончайших корешков обрывается.
Выше зоны всасывания расположена зона проведения, или зона боковых корней. На этом участке извлеченная корневыми волосками из почвы вода с минеральными солями передвигается от корня вверх по стеблю к листьям.
С первых дней жизни молодого растения корни всходов проникают на значительную глубину. Например, у клевера лугового на 30-35 см, у озимой ржи и пшеницы - 30-40 см, у овса и люпина желтого - 60-80 см, у люцерны посевной - 90-100 см. Корни взрослых растений проникают на большую глубину. Из культивируемых в Нечерноземной зоне растений наибольшую глубину проникновения корней имеют люпин многолистный (до 2,5-3 м) и люцерна посевная (до 69 м).
Корни сосны и дуба в поисках подземных вод углубляются в почве до 20-30 м и более. Глубина проникновения корневых систем растений в почву в значительной степени зависит от условий их местообитания.
Если принять во внимание, что корень обладает способностью к быстрому росту, становится очевидным, что растение не "сидит" неподвижно на одном месте. Его корни постоянно перемещаются, как бы "ощупывая" почву, проникая во все новые ее участки. Кроме хорошо развитой поглощающей поверхности корни имеют еще ряд механизмов, помогающих им "искать" пищу и воду. Слишком высокая (токсичная) концентрация почвенного раствора угнетает рост корней.
Корни не только способны находить в почве пищу, но и могут, если необходимо, превратить ее в более удобную для усвоения форму. Выделяемые ими угольная и органические кислоты способствуют использованию труднорастворимых фосфатов и других солей. Установлено также, что корни выделяют в наружную среду разнообразные ферменты. Предварительное ферментативное расщепление органических веществ помогает дальнейшему усвоению их растениями. Так, например, благодаря выделению корнями фермента фосфатазы они способны более эффективно усваивать фосфорорганические соединения, отщепляя от них минеральную форму - фосфорную кислоту.
Растения способны поглощать все элементы, встречающиеся в земной коре. Количественное соотношение поступающих в корни минеральных элементов зависит от содержания их в почве, от ее влажности и температуры и, конечно, от самого вида растений. Так, например, в одинаковых условиях корни гороха поглощают калия в 3 раза больше, чем натрия, а корни пшеницы - в 20 раз больше! Известны некоторые растения, накапливающие большое количество лития, кобальта, золота и других элементов. Растения хвоща полевого и лугового, различных видов осок и луговика дернистого (щучки) содержат много кремниевой кислоты, благодаря чему употребляются в сельской местности для чистки металлической посуды и полировки дерева.
Механизм поглощения минеральных веществ корнями растений очень важен для физиологии и в то же время весьма сложен. В клетках корня, как и в клетках зеленого листа, протекают сложные реакции.
Пища, которую доставляют корни в надземные органы растений, содержит и свободные минеральные элементы, и готовые органические вещества в виде белков, аминокислот, фосфорорганических соединений, соединений серы и железа, алкалоидов и др. Среди этих веществ есть и такие, которые создаются только в корнях и не образуются в листьях, поэтому работа листьев зависит и от состава тех веществ, которые они получают из корней. От содержания минеральных веществ в растении зависит состав протоплазмы и ее свойства, что в свою очередь определяет характер и интенсивность многих биохимических и физиологических процессов, протекающих в течение всего вегетационного периода в надземных органах.
Большую и очень ответственную работу выполняют корни по обеспечению растения водой, которая прежде всего необходима ему как составная часть организма. Зеленая масса растения содержит 80% воды и более. Вода пропитывает клеточные оболочки растения, заполняет вакуоли, придает протоплазме жидкое подвижное состояние, создает внутри клеток среду, которая наиболее благоприятна для жизненных процессов. Если растение теряет много воды, оно погибает. Только синезеленые водоросли, мхи, лишайники и некоторые другие растения могут высыхать и вновь оживать после обильных дождей. Вода необходима растениям и для построения органического вещества. Однако основная масса воды расходуется растением на транспирацию. Одно растение подсолнечника или кукурузы испаряет в среднем за вегетационный период 200-250 кг воды. С 1 га многолетнего злакового травостоя, дающего за сезон 8-10 т сена, должно испариться 5-6 тыс. т воды.
Испаряющаяся вода компенсируется новым ее поступлением из почвы, что надежно обеспечивает поддержание постоянной оптимальной температуры растения и не допускает перегрев и высыхание его листьев в жаркую погоду. Растение испаряет воду через устьица и частично через покрывающую лист кутикулу. У теневыносливых растений со слаборазвитой кутикулой внеустьичная транспирация может достигать 30% общего количества испаряемой ими воды, а у растений жаркого климата - не более 3%. Молодые листья, еще не имеющие вполне развитой кутикулы, только около половины общего количества расходуемой ими воды испаряют через устьица, а другую половину - через остальную поверхность листа.
Дерево испарило за 1 ч 500г H2О, а корневая система поглотила за это время 450г воды. Какие условия внешней среды могли вызвать указанное несовпадение количества поглощенной и испаренной воды. Как это отразится на испарении?
Библиографический список
Лесная энциклопедия: В 2-х т./Гл.ред. Воробьев Г.И.; Ред.кол.: Анучин Н.А., Атрохин В.Г., Виноградов В.Н. и др. - М.: Сов. энциклопедия, 1985.-563 с.
газета "Биология" издательского дома "Первое сентября".№ 04/2008
Лебедев С.И. Физиология растений – М.: Агропромиздат, 1988.
Тарчевский И.А. Основы фотосинтеза. – М., 1977.
5. http://www.google.ru

Нравится материал? Поддержи автора!
Ещё документы из категории биология:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ