Екологія кровосмоктучих комарів
 Зміст
Зміст
Вступ
1 Екологія кровосмокчущих комарів
2 Хвороби та умови розповсюдження хвороб преносимих комарами
2.1 Малярія
2.2 Значення комарів у передачі малярії
2.3 Умови поширення малярії
2.4 Арбовірусні хвороби
2.5 Японський енцефаліт
2.6 Західноамериканський кінський енцефаломіеліт
2.7 Східноамериканський енцефаломіеліт коней
2.8 Венесуельський енцефаломіеліт коней
2.9 Енцефаліт Сент-Луїс
2.10 Жовта лихоманка
2.11 Лихоманка денге
2.12 Лихоманка Чикунгунья
3 Комарі, переносники збудників бактеріальних хвороб
3.1 Філяриатози
3.2 Вухерериоз
3.3 Бругиоз
3.4 Туляремія
3.5 Меліоідоз чи помилковий сап
Висновок
Література
Вступ
Екологія, що виникла в надрах біології в епоху перемоги дарвінізму і розвитку історичного підходу до аналізу життєвих явищ, швидко завоювала широке визнання і стала впливати на інші біологічні науки. Вивчаючи закономірності взаємин організмів з навколишнім середовищем, екологія початку впливати на систематику тварин і рослин, різні розділи морфології (цитологію, гістологію, ембріологію й ін.). фізіологію і генетику. Виникають екологічна фізіологія, екологічна палеонтологія й інші нові напрямки. У надрах зоології розвивається екологія тварин (з підрозділами: екологія комах, екологія риб, екологія птахів і т.д.), у ботаніку — екологія рослин (екологія водоростей, екологія мохів, екологія грибів і т.д.).
В міру розвитку екології розширилися представлення про рівні організації живих систем, досліджуваних екологією: організм, популяція, вид, біоценоз, екосистема. Екологія вивчає рівні організації живого від організмів до екосистем. Біосфера не може розглядатися як рівень організації живої системи, це лише сфера життя, глобальний біотоп, у якому існують усі екосистеми, біоценози, види, популяції, організми. Але біосфера вивчається екологією також, як нею вивчаються біотопи біоценозів, місцеперебування організмів, різні фактори середовища і середовища життя. У задачу популяційної екології входить вивчення чисельності окремих видів, спалахів їхнього розмноження, закономірностей розміщення видів по території, змін популяцій за часом. Екологія видів (аутекологія) переростає в екологію співтовариств різних видів — біоценологію, а остання — у біогеоценологію, оскільки біоценози можуть існувати лише в єдності з абіотичною середовищем. Сучасна екологія включає екологію особів, екологію популяцій і екологію співтовариств. Основною проблемою екології є проблема чисельності видів. Екологія вивчає взаємодію організмів із середовищем, що обумовлює розвиток, розмноження і виживання особів, структуру і динаміку утворених ними популяцій видів, структуру і динаміку співтовариств різних видів і історично сформовані на їхній основі специфічні пристосування видів; внутрішньовидові відносини і специфічна структура виду; співтовариства популяцій різних видів, різні на різних ділянках земної поверхні, їхні взаємні пристосування, що забезпечують біогенний круговорот речовин. Важливою особливістю екології комах є необхідність вивчення особливостей життя як окремих видів, так і біоценозів у природній обстановці, вивчення комах як частини природи. До області екології відноситься вивчення явищ, у яких членистоноге виступає не як окрема особь, а як вид, вступаючи в ті чи інші взаємини з іншими видами комах, чи кліщів інших організмів, а також з фізичним середовищем. Таким чином, основним змістом екології є вивчення різноманіття взаємозв'язків організмів і середовища; закономірностей утворення і розвитку популяцій як форми існування виду, їх диференцировки і динаміки, їхньої чисельності; закономірностей формування і розвитку біоценозів як вираження взаємин організмів у конкретних умовах місцеперебування. Центральним об'єктом екології служать зв'язку, взаємини організмів із середовищем у природній обстановці і життя біоценозу в єдності з абіотичною середовищем (біогеоценоз, екологія культурних ландшафтів). Вивчення зв'язку організмів із середовищем складає теоретичну основу сучасної біології. Дані екології кровосмокчущих комах і кліщів необхідні для організації ефективних заходів щодо профілактики трансмісивних хвороб і боротьбі з паразитами людини і тварин. Екологія коштує в одному ряді з морфологією, фізіологією й іншими науками, що мають справа з усіма таксономічними підрозділами, що відбивають специфіку організму (зоологією, ботанікою, бактеріологією і більш вузькими — ентомологією, мікробіологією і т.д.). Екологія, таким чином, є одним з фундаментальних розділів біології. Використовуючи методи практично всіх біологічних наук, екологія встановлює спосіб, якої кожен вид дозволяє основну задачу, — самозбереження і поширення (Беклемішев, 1945). Екологія комах і кліщів тісно зв'язана з іншими науками. Наприклад, щоб зрозуміти шкода, заподіювана іксодовим кліщем, і запобігти його, потрібно уміти відрізняти його вид від інших. Цьому учать нас систематика і морфологія. Далі ми повинні знати спосіб життя імаго кліща і його предімагіальних фаз, місця їхній існування, характер харчування, умови, при яких вони нападають на людину, і т.д. Про все це говорить екологія кліщів. Без знань їхніх життєвих відправлень (подиху, травлення, розмноження и т.д.), про які говорить фізіологія, неможливо зрозуміти спосіб життя кліща. Приватні науки зв'язані один з одним і разом з тим зберігають свої особливості.
1 Екологія кровосмокчущих комарів
Комарі відіграють величезну роль в епідеміології ряду інфекційних і паразитарних хвороб, будучи переносниками збудників малярії, великої групи арбовірусів, гельмінтів і багатьох інших. Епідеміологічне значення кровосмокчущих комарів визначається багатьма біологічними й екологічними факторами, серед яких найбільш важливі — сприйнятливість їх до зараження, чисельність популяції, ступінь зв'язку з людиною і його житлом, активність і .тривалість життя переносника. Небезпечними переносниками збудників хвороб служать, як правило, види комарів з більшої чисельності, що харчуються переважно нa людині й активні протягом тривалого сезону.
Хвороби, збудники яких передаються комарами, найбільше-характерні для тропічних країн (жовта лихоманка, лихоманка денге, філяріатози й ін.)- Але деякі з хвороб даної групи поширені й у країнах помірного клімату (малярія, комарині енцефаліти й ін.) Зараження людини в умовах тропічного клімату можливо практично протягом усього року. Середня температура повітря тут завжди сприятлива для розвитку багатьох мікроорганізмів в організмі комарів, а виплід комарів спостерігається цілий рік. Однак чисельність комарів протягом року коливається. Так, сильні зливи в дощовий сезон нерідко змивають наявних у водоймах личинок, приводячи до зменшення чисельності комарів. У горах і передгір'ях періодичні зливи сприяють промиванню водойм, що також приводить до загибелі великої кількості личинок. У ряді районів велика кількість опадів створює додаткові водойми, сприятливі для виплоду комарів; при наявності джерел збудників захворювання це сприяє поширенню малярії й інших трансмісивних хвороб.
У сухий сезон багато невеликих водойм висихають, а водойми середньої величини зменшуються, що скорочує площу виплоду комарів і чисельність їхніх популяцій. Однак у деяких районах у цей час пересихають невеликі річечки і перетворюються в слабопроточні водойми, що служать сприятливим місцем виплоду комарів. З загального числа захворювань, у передачі збудників яких більше значення мають комарі, частина має поширення на території СРСР, а інші зустрічаються в закордонних країнах. Зростаючі з кожним роком економічні і культурні зв'язки між державами, розвиток повітряного транспорту роблять цілком ймовірної можливість завезення збудників захворювань з комарами.
2 Хвороби та умови розповсюдження хвороб преносимих комарами
2.1 Малярія
Малярія являє собою інвазионні захворювання, викликуване паразитами, що відносяться до типу найпростіших, до роду Plasmodium. Захворювання характеризується пропасними приступами, що настають через визначені проміжки часу, що зв'язано з циклом розвитку паразитів в еритроцитах.
Малярія людини викликається чотирма видами збудників: Plasmodium vivax — збудником триденної малярії, PL malariae — збудником чотириденної малярії, PL falciparum — збудником тропічної малярії і PL ovale — збудником малярії типу триденної. У Радянському Союзі місцевих випадків зараження PL ovale не установлено. Цей вид зустрічається в тропічній Африці, одиничні випадки відзначені на інших континентах.
Перераховані види збудників малярії людини розрізняються по морфологічних ознаках і характеру викликуваних ними захворювань.
В даний час маються дані про те, що люди можуть заражатися також збудниками малярії мавп — PL cynomolgi, PI, ba-stianelli, PL brasilianum. Зазначені збудники попадають в організм-людину через комарів від мавп, а в наступному — і від зараженої людини. Установлено випадки природного зараження людини PL knowlesi у природі.
Тривалість інкубаційного періоду різна і від видів (підвидів) плазмодіїв, яким заразилася людина; вона може коливатися також в окремих людей, заражених одним і тим же видом паразита, у залежності від реактивності організму. При триденній малярії тривалість інкубаційного періоду різна. При зараженні південним підвидом — PL vivax vivax інкубаційний період продовжується 10—20 днів, а при зараженні північним підвидом PL vivax hibernans — 7—10 іноді до 14 міс. При тропічній малярії інкубаційний період дорівнює 8—16 дням, при чотириденної — 3—6 нед і при малярії, викликуваної PI. ovate, 14—15 дням. При малярії, викликуваної трьома останніми видами збуджувачів, тривала інкубація не спостерігалася. Після інкубаційного періоду настає період гострих пропасних проявів малярії, що при типовому плині складається із серії пропасних приступів, що чергуються через рівні проміжки часу.
Типовий малярійний приступ включає три стадії: ознобу, жару і поту. Іноді протягом 2—3 днів приступу передують продромальні явища: загальне нездужання, головний біль, болі в кінцівках, познаблювання.
При триденній і чотириденній малярії приступ починається -: ознобу, температура в цей період доходить до 38°. При тропічній :малярії озноб виражений слабкіше, спостерігається лише невелике познаблювання, але для приступу характерні різкий головний біль, м'язові болі, нудота, блювота, можливий понос. Озноб триває від 30 хв до 2—3 ч. Озноб змінюється жаром, температура піднімається до 40—41°, загальний стан хворого різко погіршується, з'являється задишка, : можливі блювота, марення, судороги, утрата свідомості. Потім температура падає (іноді до 35,5—35°), з'являється рясний піт, слабість. При тропічній малярії піт може бути менш рясним чи відсутнє зовсім.
Типовими симптомами малярії є: анемія, що розвивається після декількох приступів, а також збільшення селезінки і печінки. Нижній край селезінки може досягати рівня пупка і навіть опускатися в малий таз.
Іноді тропічна малярія може приймати особливо злоякісне плин, така форма хвороби називається коматозною малярією, чи малярійною комою. Загальне число приступів у різних людей різко коливається, їх може бути 10—15 і більш, при чотириденній малярії звичайно більше, ніж при інших формах.
Джерелом збудників малярії людини є хвора людина (у період первинної серії приступів і рецидивів) чи паразитоносій.
Комарі заражаються на людині в той період, коли в його крові є достатня кількість гаметоцитів. На початку хвороби під час перших приступів при триденній малярії гаметоцитів мало, і можливість зараження комарів менша. При тропічній малярії гаметоцити з'являються тільки після 3—4-го приступи (через 6—10 днів після початку хвороби).
При рецидивах гаметоцити з'являються в крові в день першого приступу, а іноді і трохи раніш, тому хворий у період рецидиву значно небезпечніше, як джерело збудників, чим хворий, у перші дні хвороби. Віддалені рецидиви відрізняються великою кількістю паразитів у крові і ранній появі гаметоцитів.
Велике епідеміологічне значення має паразитоносіївство, тобто наявність паразитів у крові без явних клінічних проявів малярійної інвазії. Розрізняють первинне паразитоносіївство і паразитарні рецидиви.
У первинних паразитоносіїв паразити виявляються в крові без яких-небудь раніше виявлялися клінічних симптомів захворювання (чи до їхньої появи). Якщо такі люди володіють високої; опірністю до інвазії, то в них і надалі може не-наступити захворювання. При паразитарних рецидивах носійство настає після клінічного прояву малярії, що спостерігалося у виді гострих приступів, або могло бути подавлене застосуванням лікарських препаратів у порядку хіміопрофілактики. Паразитарні рецидиви бувають ближніми і віддаленими, і за часом збігаються з відповідними клінічними рецидивами.
Паразитоносії є схованим джерелом збудників малярії. У період великих епідемічних спалахів питома вага паразитоноссіїв як джерел інвазії досягає 30—40% і більш. Найбільшого поширення паразитоносійство досягає на наступний рік після підйому захворюваності малярією. Особливе значення має носійство збудників тропічної малярії, при якій гаметоносії є єдиною сполучною ланкою між щорічними спалахами захворювань у літньо-осінні місяці.
У природних умовах зараження людей малярією відбувається тільки за допомогою укусу комарів роду Anopheles.
Передача малярійних паразитів комарами носить сезонний характер і відбувається в теплий період року — у сезон активності комарів. Виключення складають тропічні райони, що прилягають до екватора, де зміна сезонів року виражена слабко, тут передача малярії і захворювання нею відбуваються цілий рік. Період, протягом якого можливе зараження малярією людини через укуси комарів, що заразилися плазмодіями в даному році, називається малярійним чи епідемічним сезоном. Цей період продовжується з моменту появи спорозоітів у слинних залозах комарів до відходу комарів на зимівлю.
Тривалість сезону зараження людини малярією залежить від кліматичних умов і виду комарів, і тому в різних ландшафт-але-кліматичних зонах різна. Так, наприклад, на півночі малярійний сезон продовжується місяц-півтора (частіше II-III декади липня і I-III декади серпня), при низьких літніх температурах цей період скорочується ще більше. У середній смузі СРСР малярійний сезон триває 2—3 міс, зараження людей починається в червні й у серпні, у вересні комарі ідуть на зимівлю ( що ідуть на зимівлю самки звичайно крові не п'ють). На півдні СРСР цей сезон подовжується до 5—6 міс, починаючи з травня і закінчуючи у вересні — жовтні.
В організм комара малярійні паразити попадають із кров'ю чи хворого паразитоносія (при кровосмоктанні). У шлунку (середній кишці) комара безстатеві форми паразитів гинуть, полові форми — гаметоцити проробляють подальший розвиток, дозрівають і перетворюються в зрілі полові форми — гамети. Чоловічий гаметоциг перетерплює при цьому значні морфологічні зміни. Ядро його поділяється на 4—8 частин, з цитоплазми викидається таке ж число тонких, довгих, рухливих бичів (процес ексфлагелляції). Потім бичі, що містять одне ядро, відриваються від клітки і вільно рухаються у вмісті шлунка. Кожен такий бич і є зрілою чоловічою половою кліткою — мікрогаметою.
Жіночий гаметоцит звільняється від еритроцита і перетворюється в макрогамету. Рухаючи, макрогамета зустрічає жіночу полову клітку — макрогамету, впроваджується і запліднює її. Запліднена клітка — зигота витягається і перетворюється в червоподібну,, рухливу форму — оокінету. Оокінета залишає порожнина шлунка комара, проникає в його стінку, проходить між клітками эпителия-и під зовнішньою м'язово-еластичною оболонкою округляється, перетворюючи в ооцисту. З моменту засмоктування комаром крові до утворення ооцисти, при температурі 25°, проходить близько 2 діб.
Надалі вміст цисти багаторазово поділяється, розміри її збільшуються до 50—60 мкм, у цисті утвориться велика кількість спорозоітів, що мають вид веретеноподібних кліток довжиною 14— 15 мкм і шириною в 1,5 мкм. Після дозрівання ооцисту розривається, спорозоіти попадають у порожнину тіла комара і з гемолімфою розносяться по всьому тілу, а потім вибірково накопичуються в клітках епітелію слинних залоз і в їхніх протоках.
Кількість ооцист на шлунку комара по-різному: від одиниць до 500 і більш. Число їх залежить від достатку гаметоцитів у крові людей, у яких комар ссав кров. Установлено, що при змісті в 1 куб. мм крові менш 1—2 гаметоцитів, комарі не заражаються.
Кількість спорозоітів в одній цисті може досягати декількох тисяч, а в слинних залозах комара — сотень тисяч. Зберігаються спорозоіти в слинних залозах до 1 —1,5 міс, у цей час комарі здатні заражати людей, надалі ж спорозоіти дегенерують. При температурі нижче 0° спорозоіти в комарах швидко гинуть, але при 4—10° їхня життєздатність зберігається. Зараження людини відбувається в момент уколу через слину, що містить спорозоіти (специфічна інокуляція).
Тривалість спорогонії від моменту надходження крові, що містить гаметоцитів, у шлунок комара до появи спорозоітів у слинних залозах залежить від температури середовища, у якій знаходиться комар. Тривалість спорогонії при одній і тій же температурі в різних видів плазмодіїв неоднакова. Так, наприклад, при температурі 25° спорогонія в PI. vivax продовжується 10 діб, у PL fal-ciparum — 12 діб і в PL malariae — 16 діб. Розвиток PI vivax у комарі при температурі близько 16° затягується до 55—60 діб, а при більш низькій температурі припиняється. Нижній поріг розвитку PL falciparum знаходиться близько 17—18°, для дозрівання спорозоітів цього виду потрібно більший термін, чим для PL vivax. Розвиток PI. malariae у комарах починається також при 17—18°, але продовжується ще довше, ніж у PL falciparum.
Чим вище температура навколишнього середовища, тим швидше закінчується розвиток паразитів у комарі. При більш високій температурі в комарів розвивається також велика кількість ооцист плазмодіїв, а виходить, утворюється і більша кількість спорозоітів. Самий короткий термін розвитку — при 28—30°. Але при температурі вище 30° умови для розвитку паразита знову погіршуються. У комарів, що містяться після зараження їх PL vivax при температурі 32°, розвивалися ооцисти і спорозоіти, але в слинних залозах спорозоіти не виявлялися. Можливо, висока температура перешкоджала просуванню спорозоітів у слинні залози. Якщо незабаром після ссання крові комарів помістити при 37,5°, малярійні паразити в їхньому шлунку гинуть через 2—3 ч. До низьких температур паразити в цьому випадку менш чуттєві, при температурі від 1 до 10° загибель їх у шлунку комарів наставала тільки через 2,5 дні. Якщо на стінці шлунка комара утворилися ооцисти, паразит здатний переносити низькі температури ще довше, наприклад після 7—10 діб розвитку PI. vivax при 22—24° він витримує охолодження від 1 до 10° терміном до 3 нед.
Стадії розвитку PL falciparum у комарі більш чуттєві до низьких температур. Зниження температури до 15° протягом 10—12 днів веде до загибелі паразитів. Варто мати на увазі, що при коливаннях температури протягом доби розвиток малярійної паразиту в комарі відбувається, але тільки в та годинник, коли температура перевищує нижній температурний поріг розвитку паразитів даного виду. Так, наприклад, якщо вдень температура доходить до 25°, а вночі опускається значно нижче 16°, розвиток паразитів уночі припиняється, але вони не гинуть, а з настанням сприятливої температури розвиток продовжується.
Тривалість гонотрофічного циклу в малярійних комарів, як правило, коротше, ніж тривалість спорогонії плазмодія при тій же температурі. Тому спорозоіти можуть з'явитися тільки в самок, що проробили 3—4 гонотрофічних цикли. Якщо тривалість життя комарів у даних умовах невелика, то переважна частина комарів відмирає перш, ніж у них може завершитися спорогонія і плазмодія. Коли в даній місцевості ведеться боротьба з комарами шляхом обробки приміщень стійкими інсектицидами, скорочується чисельність комарів і зменшується тривалість їхнього життя. Самки не доживають до епідеміологно небезпечного віку, тобто до - того часу, коли в них можуть з'являтися спорозоіти в слинних :залозах. Це веде до припинення циркуляції плазмодія.
Для завершення спорогонії кожен вид малярійного паразита має потребу у визначеній сумі тепла. Сума тепла є сума градусо-суток за даний відрізок часу. Градусо-сутки — це число градусів, на яке середня температура даної доби перевищує низький температурний поріг розвитку малярійного паразита даного виду, тобто температуру, нижче якої його розвиток не може відбуватися.
Розходження в тривалості спорогонії в різних видів плазмодіїв має істотне епідеміологічне значення, тому що є причиною неоднакової ураженості населення різними видами плазмодіїв в одній і тій же місцевості, а також неоднакового географічного поширення різних видів плазмодіїв. При однаковій температурі найменша кількість часу потрібно для розвитку спорозоітів PI. vivax, для розвитку спорозоітів PL falciparum часу потрібно більше, а для спорозоітів PL malariae — ще більше. При одночасному зараженні комарів плазмодіями різних видів можливість передачі збудників триденної малярії настає раніш, ніж збудників інших видів. Унаслідок цього в одній і тій же місцевості за інших рівних умов найбільше поширення має триденна малярія, потім тропічна і менш усіх поширена чотириденна малярія.
Неоднаковий нижній поріг розвитку в комарі і різній швидкості спорогонії в різних видів плазмодіїв складають основну причину того, що далі всього на північ поширена триденна малярія, трохи південніше — тропічна, і чотириденна малярія зустрічається тільки в південних районах. Задовго до відкриття збудника малярії і її переносників епідеміологи знали, що північна границя малярії в Європі збігається з липневою ізотермою +16°. В даний час відоме понад 50 найбільш важливі види переносників, а при сприятливих обставинах ще близько 60 видів може утягувати в процес передачі плазмодіїв. Інтенсивність поширення малярії за інших рівних умов зростає з підвищенням середніх температур, від яких залежать тривалість сезону передачі і чисельність переносників, тому ефективні переносники в одних країнах можуть не грати цієї ролі в інші;. Поширення малярії не збігається з границями поширення переносників даного захворювання, що обумовлюється наступним: розвиток комарів у воді починається при температурі 10°, у той час як розвиток плазмодіїв у їхньому організмі відбувається при температурі не нижче 15—16°, але і при зазначеній температурі розвиток плазмодіїв відбувається дуже повільно. Тому границя поширення кома-рів-анофелес доходить до 65° північ. ш. Місцеві випадки малярії на цій широті зустрічаються вкрай рідко.
2.2 Значення комарів у передачі малярії
Значення того чи іншого чи виду підвиду комарів у передачі малярії залежить від ряду умов. На підставі досвіду численних досліджень роль кожного виду комарів у передачі малярії зараз оцінюється з погляду п'яти факторів, що визначають його епідеміологічну ефективність.
1. Сприйнятливість даного виду комарів до визначеного виду малярійного паразита, іншими словами, специфічність взаємин між паразитом і членистоногим-хозяином. Визначається сприйнятливість емпірично по спорозоітному чи ооцистовому індексах комарів, що розкриваються, або (що більш точно і доказово) при постановці досвідів по заражуваності комарів на хворій людині. Багато комарів Anopheles, що живуть на території СРСР, сприйнятливі до Plasmodium vivax, однак порівняльних досвідів, що показують на великому матеріалі й в однорідних умовах кількісні розходження в заражуваності різних видів, не проводилося. Це питання повинне стати одним з найважливіших у майбутніх дослідженнях. Були виявлені значні розходження в сприйнятливості різних видів до PI. falciparum. Наприклад, An. hyrcanus виявився дуже слабко сприйнятливим видом до цього паразита на відміну від An. superpictus і An. pulcherrimus. Про розходження в сприйнятливості різних видів до PI. malariae поки відомо мало.
2. Контакт комарів з людиною. Цілком природно, що для успішної передачі малярії комар повинний бути агресивний стосовно людини і мати можливість для нападу на людей. Це залежить у першу чергу від екології і поводження самих комарів, а також від особливостей побуту населення. Добре відомий поділ комарів на ендофильних і екзофіильних. Екзофільні комарі предпочитають для днювань природні біотопи. Звичайно ендофіли більш небезпечні в епідеміологічному відношенні через великий контакт із людиною, але є і екзофіли, що є дуже ефективними переносниками. Іноді в різних умовах той самий вид поводиться по-різному.
Важливо також поділ комарів на екзофагів і ендофагів. Екзофаги нападають на людей поза приміщеннями, ендофаги — усередині будинків, особливо під час сну. Комарі ряду видів можуть нападати як усередині приміщень, так і зовні. Необов'язково, щоб екзофаг був ендофилом; наприклад, An. plumbeus може масами нападати на людей усередині будинків, а потім залишати їх і вибирати днювання в природі. Велике значення мають і поведінкові особливості комарів, зв'язані з розміром видобутку. Наприклад, для таких видів, як An. superpictus і An. pulcherrimus, більш притягальні великі чи тварини скупчення людей і тварин. При цьому комарі можуть літати здалеку в чи селища тваринницькі ферми, ігноруючи шляху, що зустрічаються їм на, окремих людей. На противагу їм існують і мисливці за дрібним розсіяним видобутком: дрібними ссавцями, птахами і т.п.
Контакт людини з комарами визначається різними способами. Отут і порівняльні вилови на людині і тваринах усередині і поза приміщеннями, усередині і поза селищами, установка різних пасток на вліт і виліт комарів, визначення приналежності крові зі шлунків ситих комарів за допомогою реакції преципітації. Для зменшення контакту комарів з людиною в СРСР широко практикувалася так називана зоопрофілактика, тобто відволікання комарів худобою. Для цього тваринницькі ферми встановлювалися між місцем виплоду комарів і селищем. Деякі види не можуть бути переносчиками малярії просто через те, що придатні для них місця виплоду знаходяться далеко від селищ, і, таким чином, контакту із людиною вони не мають.
3. Чисельність комарів. За інших рівних умов вона є важливим чинником, що впливає на епідеміологічну ефективність комарів. При більш високій чисельності збільшується кількість самок, що нападають на людину, і, як наслідок, інтенсивність передачі. Однак це справедливо тільки в тому випадку, якщо інші фактори незмінні. Особливо помітні розходження чисельності різних видів. Наприклад, An. messeae у СРСР чи на півночі Європи звичайно дуже рясний, але є поганим переносником через інші фактори. Африканський An. gambiae, навпаки, навіть при низької чисельності здатний підтримувати інтенсивну передачу малярії. Чисельність залежить від багатьох факторів: кількості місць виплоду, прокормувачів, виживаності, на которую впливають погода і хижаки, і ін. Протягом сезону вона міняється. Найбільш інтесивна передача малярії звичайно відбувається під час її піків. Звичайно маляріології судять про чисельність виду в той чи інший момент по показниках відносної чисельності, тобто по числу особ, зілишаємих за одиницю часу або на одиницю поверхні. Для спеціальних цілей іноді приходиться встановлювати абсолютну чисельність популяції або її частини на конкретній території.
4. Виживаність комарів до фізіологічно небезпечного віку. Після кровосмоктання на хворому самка комара повинна прожити досить тривалий час, щоб малярійний паразит пройшов необхідний цикл розвитку і зробив комара заразним. Тривалість циклу спорогонії паразита залежить від температури і легко визначається за допомогою методів Мошковского чи Оганова — Раєвського. Протягом циклу спорогонії самка проходить трохи гонотрофічених циклів, що визначають її фізіологічний вік. Звичайно, щоб стати эпідеміологічно чи потенційно небезпечної, самка комара повинна проробити два чи більш гонотрофічних цикли. Їхня тривалість залежить від температури і вологості повітря. На кожнім гонотрофічному циклі гине визначена кількість самок, особливо при відкладці яєць. Таким чином, чим нижче природна смертність комарів, тим вище чи небезпека інтенсивність передачі малярії і навпаки. Смертність комарів залежить від багатьох факторів, але особливо великий вплив на неї роблять погодні умови.
5. Температура повітря. Цей природний фактор впливає на всі попередні. Температура насамперед визначає видовий склад комарів у даній місцевості. Від її залежать тривалість сезону передачі, швидкість циклу спорогонії, швидкість гонотрофічних циклів, чисельність і активність нападу комарів « їхня виживаність.
На території СРСР температурний фактор особливо важливий.
У холодних, самих північних районах узагалі немає комарів. Трохи -південніше, де вони є, тепла для завершення навіть одного циклу спорогонії в комарі недостатньо. Просуваючи на південь, потенційну небезпеку виникнення малярії усе більш збільшується, оскільки" зростають число видів комарів, швидкість циклу спорогонії, і, як, наслідок, число послідовних циклів спорогонії за сезон.
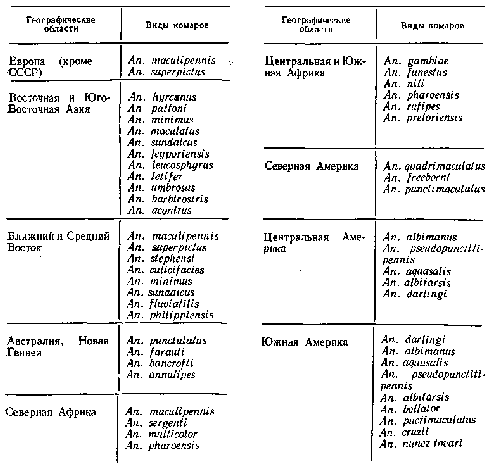
Отже, резюмуючи усе викладене, можна укласти, що в даний час потенційно найбільш небезпечними видами в нашій країні1 є An. sacharovi (кавказький), An. superpictus і An. pulcherri-mus. Менш ефективними переносниками служать An. maculipennis, An. sacharovi (середньоазіатський), An. I. atroparvus і An. messeae. Інші комарі (An. hyrcanus, An. melanoon, An. plumbeus, An. cla-viger) другорядні й епізодичні переносники малярії. В Індії з 42 видів, що там зустрічаються, Anopheles тільки 7 відносять до важливих переносників малярії. У Бразилії з 50 видів Anopheles 4 вважаються основними переносниками і 5 — другорядними.
У загальному розходження в ступені заражуваності між видами частіше невелико, одні види переносників заражаються приблизно в 2— 3 рази краще інших (якщо в An. maculipennis чи An. superpictus заражаються PI. vivax 80—90% особ, то в An. hyrcanus — 30—40%). Рідше ці розходження можуть бути дуже значні: так, An. hyrcanus-заражається збудниками тропічної малярії раз у 30 рідше, ніж А п. superpictus. Нерідке зараження комарів настає тільки при повторному кровосмоктанні. У різних географічних областях земної кулі лише відносно невелика кількість видів Anopheles може бути віднесене до важливого ( чифактично потенційно) переносникам малярії (табл. 1.1).
До малярії сприйнятливі практично всі люди. Клінічний плин захворювання може бути різним, що залежить від стану організму, що неоднаково реагує на зараження навіть одним і тим же штамом плазмодія.
Імунітет при малярії строго специфічний і виникає тільки до визначеного виду і штаму збудника. Завдяки цьому чи хворій паразитоносій стійкий до повторного зараження ідентичним штамом, але легко заражається іншим чи штамом видом плазмодія.
2.3 Умови поширення малярії
Поширення малярії, як і інших інфекційних захворювань, у тій чи іншій місцевості можливо тільки при наявності джерел збудників, комарів — переносників збудників і сприйнятливих до малярії людей. Крім цих умов для поширення малярії потрібні відповідні температури, необхідні для завершення розвитку плазмодіїв (спорозоітів) в організмі комарів. Температурний мінімум, при якому відбувається розвиток найбільше «холодолюбивого» виду плазмодіїв — PL vivax, + 16°. При більш низьких літніх ізотермах (нижче 16°) комарі не доживають до кінця спорогонії.
Територія можливого поширення малярії в східній півкулі знаходиться між 63° північ. ш. і 30° півд.ш., а в західній півкулі — між 40° північ. ш. і 40° півд.ш. Температурний фактор обмежує поширення малярії й у горах, наприклад, у Середній Європі вона відсутній на висоті більш 500 м над рів. м., в Італії — понад 1000 м, у тропічних областях — вище 2000 м, але в деяких районах малярія спостерігається і на великих висотах (у Мехіко — на висоті 2200 м, у Ла-Пасі — 3600 м).
Інтенсивність поширення малярії залежить від ряду умов, зокрема, соціально-економічних, кліматичних, ландшафтних. Вона залежить також від екологічних особливостей переносників і можливостей для їхній виплоду. Триденна малярія — найбільш розповсюджена форма, що зустрічається як у північних широтах, так і в тропічному поясі. Триденна малярія з тривалою інкубацією переважала в північній і середній смузі європейської частини СРСР, у Сибіру, на півночі Казахстану, у деяких країнах Європи. У південних областях СРСР переважала триденна малярія з короткою інкубацією, що спостерігається звичайно приблизно з 44° північ. ш. У Закавказзі і республіках Середньої Азії з усіх випадків триденної малярії тільки 10% захворювань починалися з тривалою інкубацією.
Ovale-малярія в одиничних випадках реєструється на всіх континентах. Найбільше часто вона зустрічається в Східній Африці, причому в дітей ця форма буває в 3—5 разів частіше, ніж триденна малярія, викликувана PL vivax (у дорослих — рідше). У Західній Африці PI. ovale знаходили в 10% дітей — паразитоносіїв. У СРСР доведених місцевих випадків оvale - малярії не виявлене.
Тропічна малярія поширена переважно в тропіках і субтропіках.
Чотириденна малярія реєструється значно рідше, ніж триденні чи тропічна, вогнища її зустрічаються в Африці, Індії, Індонезії, Центральній і Південній Америці. Комарі роду Anopheles можуть передавати і збудників вірусних хвороб. Так, зі спонтанно заражених комарів різних видів цього роду в Північній і Південній Америці, Африці, Австралії, Індії, Японії, Чехословакії і на Ямайці виділені 24 вірусу різних груп. Комарі An. funestus і An. gambiae були найголовнішими переносниками збудника лихоманки о'ньонг-ньонг у Східній Африці в 1959 р., коли зареєстрували понад 5 млн випадки цього захворювання. При цьому виділено по 10 штамів вірусу з кожного виду переносника. На південно-сході США в 1960—1962 р. з An. crucians був виділений новий вірус групи Буньямвара — вірус Tensaw (Chamberlain, 1963).
Таблиця 2.1 – Основні види переносників збудників малярії
2.4 Арбовірусні хвороби
До групи арбовірусів (від скорочених: слів arthropod-borne, тобто стерпні членистоногими) відносяться віруси людини і тварин, що розмножуються в організмі кровосмокчущих членистоногих і передаються хребетним через слину чи комахами кліщами при кровосмоктанні. З більш 300 відомих арбовірусів 72% передають комарі. Загальне число арбовірусів, поданим обстеження різних континентів, істотно зменшується в міру просування від екваторіального кліматичного пояса до помірного. Ці тенденції чітко виявляються у випадку арбовірусів,, переданих комарами (у групі вірусів, переданих кліщами,, цих тенденцій немає). Всі арбовірусні хвороби є природно-осередковими, і їхнє поширення визначається набагато великим числом природних факторів, чим антропонозних нетрансміссивних вірусних хвороб (кору, грипу, віспи й ін.).
Поширення арбовірусів залежить від наявності специфічних: переносників, сприйнятливої хребетної тварин, суми тепла,, опадів і інших факторів. Одні віруси передаються строго специфічними видами і родами комарів, інші — різноманітними представниками багатьох пологів. Наприклад, вірус західного енцефаломіеліту коней у природних умовах виділений не менш чим від 20 виглядів-комарів пологів Culex, Aedes, Anopheles, Culiseta і Psorophora. А вірус лихоманки денге передають комарі лише одного роду Aedes. Однак-відсутність вузької специфічності у фізіологічному плані не означає, що той чи інший вірус у природі переносять усі комарі, залученні в цикл його циркуляції. Незважаючи на те що вірус західного енцефаломіеліту коней виділений від багатьох видів комарів, його циркуляція на більшій частині ареалу забезпечується одним видом комара — Culex: tarsalis. Рівною мірою це стосується й інших арбовірусів, виділених від декількох видів комарів. Вірус жовтої лихоманки в природі виділений тільки від комарів Aedes і Haemagogus, хоча в його передача легко здійснювалася багатьма видами інших пологів комарів.
Усе це свідчить про те, що далеко не кожний з потенційних переносників є таким фактично. Причина ц-явища полягає в обмежуючому дії факторів екологічного порядку. Одним з важливих екологічних факторів, що визначають циркуляцію арбовірусів у природі, є специфіка зв'язків переносників із хребетними хазяїнами арбовірусів. Трофічні зв'язки комарів із хребетними хазяїнами арбовірусів визначають основні цикли циркуляції. Дуже багато видів Culex і Mansonia харчуються в основному на птахах; Aedes, Anopheles і Culiseta — на ссавец. У групі А арбовірусів 6 вірусів зв'язані з комарами Culex і птахами, а 10 — з комарами Aedes, Anopheles і Haemagogus і ссавцями. У групі Б арбовірусів 11 вірусів циркулюють у системі птах — комарі Cutex чи Mansonia — птах, а 10 вірусів — у системі ссавці — комарі Aedes — ссавці.
Майже всі арбовіруси, що викликають значні епідемії, є вірусами ссавців і циркулюють за допомогою комарів Aedes (жовта лихоманка, лихоманка денге й ін.). Частина комарів нападає на рептилій, передаючи їм арбовіруси інших хребетних. Так, вірус західного енцефаломіеліту коней може передаватися комарами зміям. У зміях цей вірус перезимовує, а навесні включається в звичайне коло циркуляції за допомогою комарів.
Переважна більшість арбовірусів, стерпних комарами, має тропічне походження. Цим порозумівається їхня підвищена вимогливість до температурних умов. Відомо, наприклад, що вірус жовтої лихоманки в тілі комара Aedes aegypti при температурі 37° досягає дози, що інфікує, через 4 дні, при° 36 - 6, при° 25 - 8, при° 23 - 11, при° 21 - 18 днів, а при° 18 вірус у комарах не виявляється навіть через 30 днів. Аналогічні результати у відношенні цього вірусу отримані в експериментах з Haemagogus. Температурний режим, безсумнівно, є найважливішим чинником, що обмежує ареал жовтої лихоманки, тому що переносники вірусу поширені значно ширше, ніж сам вірус.
Аналізуючи географічне поширення деяких арбовірусів, стерпних комарами, можна помітити, що в умовах континентального клімату середніх широт північної півкулі північна границя їхнього можливого поширення визначається тривалістю періоду з температурами вище 10°. Даний період, як показали дослідження Д.К. Львова й А.Д. Лебедєва (1974), повинний складати не менш 210 днів. Переважна більшість арбовірусів, стерпних комарами, не виходить за межі 30° північної і південної широт. Значне число відомих арбовірусів поширене в межах одного чи двох-трьох материків, якщо між ними маються материкові зв'язки (зокрема, Сівши. і Юж. Америка). Лише жовта лихоманка і лихоманка денге поширені на різних материках, що не мають континентальних зв'язків, чи на материках і вилучених океанічних островах. Поширення цих вірусів зв'язано з міжконтинентальними переміщеннями чи людини птахами-мігрантами.
Важливу в епідеміологічному відношенні особливість представляють так називані викарируючі віруси, що ніколи не зустрічаються разом. Наприклад, викарируючими близькородиннми вірусами є вірус західного енцефаломіеліту коней, розповсюджений в Америці, і Синдбіс, що зустрічається в Африці й Азії; Сент-Луїс в Америці і віруси Західного Нілу і японського енцефаліту в Африці, Європі й Азії.
Кількість арбовірусів знаходиться в прямої залежності від видового достатку теплокровних хазяїнів і комарів, без яких існування арбовірусів у природі неможливо.
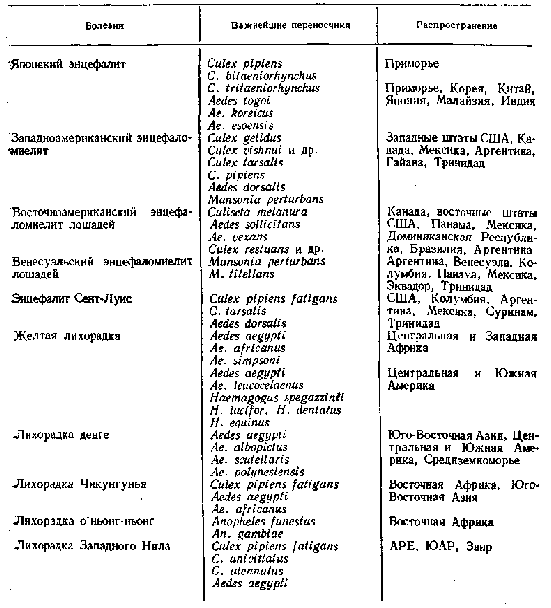
У табл. 1.2 приведені арбовірусні хвороби, збудники яких чи переважно винятково передаються комарами. Однак відомо багато арбовірусів, що передаються іншими членистоногими, але в передачі їх беруть участь і комарів. Як приклад можна привести виділення вірусу лимфоцитарного хориоменінгіту з комарів Aedes communis, Ae. dianteus, Ae. cantans у районах Західної України; вірусу кліщового енцефаліту з комарів Aedes до ін.Роль комарів як переносників визначається можливістю інтенсивного розмноження вірусу в організмі комахи і нагромадження вірусу в слинних залозах у кількості, достатньому для інфікування хребетного хазяїна. При наявності комарів-переносників, що харчуються як на диких тваринах, сприйнятливих до інфекції, так :і на людях, вірус може іррадирувати із природних вогнищ у населену зону. Прикладом може служити спалах жовтої лихоманки в Ефіопії в 1960—1962 р., коли занедужало понад 15000 чоловік при летальності 85%. Сполучною ланкою між інфікованими тваринами (мавпами) і людиною служив, очевидно, комар Ае. simpsoni, що живе як у лісах, так і в селищах, нападаючи і на мавп і на людей (Гуцевич, Мончадский, Штакельберг, 1970).Японський енцефаліт — захворювання, викликуване вірусом,, характеризується важким клінічним плином з переважною поразкою центральної нервової системи й у 60% випадків закінчується летально. Уперше це захворювання було описано в 1924 р. у Японії (відкіля і відбулася назва хвороби), коли під час великого епідемічного спалаху летальність серед захворілих досягла 60%. У 1938 р. захворювання уперше виявлене на території СРСР, у Приморському краї.
Інкубаційний період коливається від 4 до 14 днів. Поразка центральної нервової системи характеризується вираженими змінами мозкового стовбура і базальних ядер головного мозку. У видужалих на відміну від хворих кліщовим енцефалітом залишкових явищ у виді паралічів звичайно не спостерігається. Після перенесеного захворювання розвивається тривалий імунітет; повторних захворювань майже не буває.
2.5 Японський енцефаліт
Японський енцефаліт — природно-осередкове захворювання. Резервуарами вірусу в природі є дикі птахи (горобці, фазани, чаплі й ін.). В організмі птахів збудник викликає частіше безсимптомну інфекцію і зберігається в міжепідемічний період. У крові-птахів виявляються специфічні антитіла до вірусу японського енцефаліту.
Таблиця 2.2
Арбовірусні хвороби, збудники яких передаються комарами
Специфічними переносниками вірусу японського енцефаліту є комарі пологів Culex і Aedes: Culex pipiens, С. triiaeniarhyn-chus, C. bitaeniarhyhchus, Aedes togoi, Ae. koreicus на території СРСР і в суміжних країнах. Крім того, у природних вогнищах закордонних країн епідеміологічне значення мають комарі Ae. eso-ensis (Китай, Японія), Culex gelidus (Малайзия, Індія) і деякі Інші.
У період активності комарі переносять вірус від птахів до людини і теплокровних тварин.
Тривалість циркуляції вірусу в організмі теплокровн-тварини і людини обчислюється коротким терміном, очевидно, декількома днями. Тому людина і тварини — лише тимчасові носії вірусу, однак вони відіграють відому роль у зараженні нових партій комарів.
Механізм передачі вірусу комарами — специфічна інокуляція. Збудник проникає в організм людини при укусі комара і поширюється з кров'ю по всьому організмі, розмножуючи переважно в силу нейротропності в центральній нервовій системі. Заражені вірусом енцефаліту комарі є довічними носіями вірусу, що зберігається при їхній перезимовки.
На розвиток вірусу енцефаліту в організмі комара впливає температура навколишнього середовища. При температурі нижче 20° розвиток вірусу в тілі комара загальмовується, що обумовлює поширення природних вогнищ даного захворювання в місцевостях з теплим кліматом і досить високою вологістю. Трансовариально вірус у комарів не передається.
Для розвитку зазначених комарів велику роль грає середня температура і вологість повітря в літні місяці. Оптимальні умови для прискореного циклу розмноження комарів створюють у період жаркого літа з великою кількістю сонячних днів при середній температурі повітря 22—25° і високої відносної вологості до 80—90%.
На Далекому Сході в жаркі літні місяці нерідкі розливи рік і повені, викликувані інтенсивним таненням снігу в горах і заливними дощами, що веде до розвитку великих заболоченостей, що забезпечують масовий виплід переносників. Липень і серпень характеризуються найвищим температурним режимом, що сприяє росту чисельності і посиленню активності переносників і нагромадженню вірусу в їхньому організмі. Цей відрізок часу можна вважати предепідемічним періодом. З другої половини серпня до середини жовтня можливі захворювання японським енцефалітом серед населення.
Японський енцефаліт широко розповсюджений у країнах Східної і Південно-Східної Азії: у Кореї, Китаї, Японії, Малайзії, Індонезії, на Філіппінських островах, в Індії і на островах Індійського океану. У СРСР природні вогнища японського енцефаліту маються в південом Примор'я.
2.6 Західноамериканський кінський енцефаломіеліт
Західноамериканський кінський енцефаломіеліт — гостре вірусне захворювання з переважною поразкою центральної нервової системи. Хвороба характеризується високою температурою (до 40—41°) протягом 7—10 днів і нерідко супроводжується (у 10—15% випадків) паралічами. Летальність серед дорослих не перевищує 7—8%, серед дітей вона досягає 20%.
Резервуарами збудника в природі є птахи (дикі качки, гусаки, фазани, горобці й ін.), від яких комарі і гамазові кліщі передають вірус коням, мулам, дрібним гризунам і людині. Спонтанне вірусоносіївство виявлено також у змій і жаб. Людина частіше заражається в антропургічн их вогнищах, де джерелом збудника є домашні і синантропні тварини.
Найбільш частими переносниками є комарі С. tarsalis, С. pipiens, Ae. dorsalis, Ae. vexans, Mansonia perturbans і інші. Переносити вірус людям і коням від основного джерела збудника захворювання (диких птахів) можуть лише види, що часто нападають на ссавців і птахів. С. tarsalis охоче нападає на корів і коней, але часто харчується також на птахах, кров яких виявляється в 50—98% особ. У комарів Mansonia perturbans пташина кров виявлена в 27%, а в Ae. vexans у 5% випадків.
Передача збудника комарами здійснюється за допомогою специфічної інокуляції. Вірус, потрапивши в кишечник комара, розмножується і з гемолімфою поширюється по всьому організмі. Протягом першого тижня після кровосмоктання на хворому вірус виявляється в слинних залозах комара, де поряд з кишечником знаходиться найбільш кількість вірусу.
Захворювання реєструються звичайно із середини липня до вересня, що відповідає найбільшої активності комарів. Воно зустрічається в східних штатах США і Канаді.
2.7 Східноамериканський енцефаломіеліт коней
Східноамериканський енцефаломіеліт коней — вірусне епізоотичне захворювання коней і птахів. Через укуси кровосмокчущих членистоногих вірус може потрапити в організм людини і викликати в нього важке захворювання з переважною поразкою центральної нервової системи. Хвороба характеризується високою температурою, головним болем, судорогами і паралічами, продовжується від декількох днів до трьох тижнів. У 60% випадків спостерігаються залишкові явища після хвороби — паралічі, розумові розлади. Летальність досягає 65%.
Основним резервуаром вірусу в природі є птахи, особливо дрібні, у яких спостерігається більш тривала вірусемія з більш високими титрами вірусу, чим у великих птахів. Природними вогнищами східноамериканського енцефаломіеліту на сході США служать прісноводні болота, де протягом зими вірус зберігається в гризунах і інших тваринах.
Основним переносником у циркуляції вірусу серед диких птахів є комар Culiseta melanura, на частку якого приходиться більше половини виділених штамів вірусу східноамериканського енцефаломіеліту. Вірус неодноразово виділявся і від інших видів комарів (М. perturbans, Ae. vexans, Ae. sollicitans, Ae. mitchellae, An. crucians, С. restuans, С. salinarius). Відомі також факти виділення вірусу від мокреців, гамазових кліщів (Dermanyssus gallinae) уший (Eumenacanthus straminaeus).
Під час епідемій у передачу вірусу утягують Ae. aegypti, Ae. triseratus, Psoropha discolor. Вони можуть заражатися, харчуючи кров'ю з дуже невеликим змістом вірусу і передавати його з 50— 90%-ний ефективністю. Комарі Culex fatigans, С. salinarius R An. quadrimaculatus заражаються в невеликому відсотку випадків і та лише при високій концентрації вірусу в крові і передають його менш чим з 5%-ний ефективністю. Culex tarsalis має дуже низький поріг зараженості і здатний передавати вірус від птаха до птаха. Ae. sollicitans може передавати вірус від коня до коня, якщо титр вірусу в крові донора трохи вище рівня, характерного для даного виду. Звідси стає зрозумілим, що надзвичайно високий рівень чисельності комарів, що збігає з активністю вірусу, ще не визначає рівня епізоотії. Тут важливим є співвідношення комарів, чисельність потенційних переносників вірусу.
Захворювання серед людей реєструється звичайно протягом серпня — вересня. Механізм передачі збудника — і специфічна; інокуляція. Комарі можуть служити не тільки специфічними, але і механічними переносниками вірусу. Мухи Stomoxys calcitrans можуть передавати вірус протягом 4 ч після зараження. східноамериканський енцефаломіеліт розповсюджений на території східних штатів США, у Канаді, Панамі, Бразилії й Аргентині, частіше в сільських місцевостях.
2.8 Венесуельський енцефаломіеліт коней
Вірус даної хвороби уражає людей і коней у ряді країн Південної Америки-(Венесуела, Колумбія, Еквадор, у Тринідад). Захворювання характеризується сильним головним болем, болем в очних яблуках, м'язах, особливо ікроножних, шийних і поперекових. Смертельні ісходи рідкі.
Резервуар вірусу в природі не знайдений. Захворювання людей спостерігаються, як правило, у весняно-літній період (березень — липень) і зустрічаються переважно в сільській місцевості.
Основними переносниками вірусу венесуельського енцефаломіеліту є комарі Mansonia titellans, Ae. taeniarhynchus, що передають вірус шляхом специфічної інокуляції.
2.9 Енцефаліт Сент-Луїс
Енцефаліт Сент-Луїс відноситься до групи американських комариних енцефалітів; у 20% випадків супроводжується летальністю. Резервуаром вірусу є птахи, від яких він за допомогою комарів передається людям і коням. Захворювання людей спостерігаються наприкінці літа і восени (серпень — вересень).
Переносниками збудника енцефаліту Сент-Луїс служать комарі пологів Culex і Aedes: С. pipiens, С. tarsalis, С. fatigans, Ae. dorsa-lis і ін. Крім комарів вірус можуть передавати також паразитуючі на птахах гамазові кліщі. Захворювання поширене в США, Південній Америці (у Сурінаме і на о-ві Тринідад).
2.10 Жовта лихоманка
Жовта лихоманка є гострим пропасної захворюванням вірусної етиології. Захворювання розвивається після 4—7-денні інкубації і характеризується високою температурою, поразкою печінки і бруньок, важким загальним станом, крововиливами в порожнині рота, шлунка і кишечнику, жовтяницею. Летальність коливається від 5; до 80%, у середньому вона дорівнює 25—30%.
Жовта лихоманка — облігатно-трансміссивне захворювання з природної очаговістю. Розрізняють дві епідеміологічні форми жовтої лихоманки: лихоманку джунглів і лихоманку населених пунктів. Перша форма зв'язана з природними вогнищами, у яких резервуаром вірусу є мавпи і деякі сумчасті (опосум) f а переносниками — різні види комарів пологів Aedes і Наета-gogus. При лихоманці населених пунктів джерелом збудника захворювання є людина, а переносником — синантропний комар-Aedes aegypti.
Природні й ендемічні вогнища жовтої лихоманки розташовані у вологих тропічних лісах, переважно в басейнах рік: Амазонки і Конго.
У Південній Америці (Бразилія, Колумбія) основними переносниками вірусу в природі є комарі Haemagogus spegazzinii і Aedes leucocelaenus, а в Центральній Америці — Haemagogus lucifer, Н. spegazzinii, H. dentatus, H. equinus. У Центральній Африці природні вогнища жовтої лихоманки підтримуються за рахунок мавп, серед яких вірус передається комарами роду Aedes переважно Ае. аїґісапиз. Важливим фактором є здатність Ае. africanus переживати сухий сезон у кронах лісу і цим забезпечувати підтримка діючого вогнища жовтої лихоманки.
Заражаються в природних вогнищах найчастіше лісоруби, робітники, що прокладають дорогу в лісі, військовослужбовці. Отриманий людиною вірус надалі поширюється серед населення селищ антропофільним комаром Ае. aegypti.
Комар Ае. aegypti тісно зв'язаний з житлом людини; виплужується він у найрізноманітніших умістищах для води поблизу приміщень і в самих будинках, у декоративних водоймах, консервних банках, різних горщиках. У цих типах вогнищ хвора людина є: єдиним джерелом збудника хвороби, тобто тут жовта лихоманка є типовим антропонозом.
Вірус жовтої лихоманки зберігається в організмі комара довічно, але трансовариально не передається. Комар здобуває здатність заражати через 12 дн. після кровосмоктання на хворому, що морозить, якщо температура повітря за цей час не падає нижче°25 . Чим вище температура середовища, тим коротше термін інкубації вірусу в комарі (до 4 днів при 37°) і навпаки. При спаді температури нижче 20° заражений комар утрачає здатність передавати вірус, однак при наступному підвищенні температури він знову стає здатним заражати людей і тварин. Це порозумівається швидк-розмноженням вірусу жовтої лихоманки у всіх тканинах комара (комар при цьому не хворіє) при температурі 25—30°, що забезпечує можливість переносу збудника здоровій людині при наступному укусі.
Температурний фактор обмежує поширення жовтої лихоманки жаркими країнами. У країнах з помірним кліматом, де температура повітря тільки на короткий термін піднімається до 25°, можливий спорадичний замет вірусу, і завезені туди заражені комарі можуть передавати збудника захворювання. Замет комара в різні райони земної кулі сучасним повітряної і морським транспортом приводить до появи спалахів жовтої лихоманки в .багатьох місцевостях з теплим кліматом.
Сприйнятливі до вірусу жовтої лихоманки люди усіх віків, але в ендемічних вогнищах у зв'язку з імунологічним прошарком .доросле населення хворіє рідше, ніж діти. У природних вогнищах жовтої лихоманки джунглів занедужують переважно дорослі, тому що вони частіше працюють у джунглях.
Хоча найбільш великі епідемії відзначалися звичайно в тропичес-.ких районах, але спалаху жовтої лихоманки при заметі збудників зараженими комарами виникали майже усюди, де малися переносники, тобто від 42° північ. ш. до 40° півд.ш. в обох півкулях землі. Так, неодноразово жовта лихоманка заносилася в Іспанію, Португалію, на південь Франції й Італії. У 1857 році під час великої епідемії в Лісабоні занедужало 19000 і вмерло 7000 чіл.
Загальне число померлих від жовтої лихоманки, за даними зареєстрованих епідемій, складає більш 3 млн. чіл. Відомо, наприклад, що при будівництві Панамського каналу ця хвороба віднесла в могилу понад 10000 чіл.
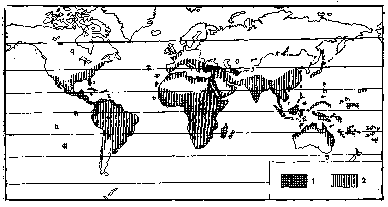
В даний час жовта лихоманка поширена в джунглях Західної, Центральної і Східної Африки і Південної Америки (Бразилія, Венесуела, Колумбія, Еквадор, Панама, Нікарагуа, Коста-Ріка, Перу, Болівія, острови Карибського моря, північно-східна Аргентина) (мал. 2.1).
Малюнок 2.1 – Географічне поширення жовтої лихоманки: 1 — ендемічні райони; 2 — райони, де виявлені антитіла до вірусу жовтої лихоманки; 3 — ареал переносників за межами ареалу жовтої лихоманки
2.11 Лихоманка денге
Лихоманка денге — осередкові-осередкову-осередкова-осередковий-природно-осередкове трансміссивне захворювання, викликуване вірусом, що передається комарами роду Aed.es. Хвороба розвивається після 4—денн-7-денні інкубації і характеризуеться різкими головними болями і болями в суглобах, м'язах, почервонінням шкіри, високою температурою і висипкою, що з'являється на 3—5-йг день захворювання. Через 5—7 днів лихоманка й інші симптоми хвороби зникають і хворий видужує. Летальність дуже незначна.
Основним специфічним переносником вірусу лихоманки денге є комар Ае. aegypti, що заражається від людини в інкубаційному періоді (за 6—18 ч до початку хвороби) і протягом перших трьох днів захворювання. Комарі починають передавати збудника через 11—14 днів після кровосмоктання на хворому і можуть зберігати його в організмі довічно. Наступному поколінню комар-вірус не передається. Комарі Ае. aegypti містять звичайно значна кількість вірусу у своєму організмі, унаслідок чого достатн-укусів 1—2 заражених комарів, щоб заразити людини. Вірус активний в організмі комарів при температурі 18° і вище.
Крім Ае. aegypti переносниками вірусу лихоманки денге служать Ае. albopictus, Ae. scutellaris, Ae. polinesiensis. Усі вони відносяться до підроду Stegomyia, розповсюдженому в тропічних районах. Ці комарі здійснюють перенос вірусу серед деревних тваринних тропіків: мавп, лемурів, білок. Ае. albopictus може передавати збудника від тварин людині. Ае. aegypti передає вірус лихоманки денге тільки в епідемічних антропонозних вогнищах.
Захворювання лихоманкою денге поширені в Східній і Південно-Східній Азії (Корея, Китай, Індокитайський півострів, Індія, Індонезія, Філіппіни, Японія). У Японії під час другої світової війни щорічно спалахували епідемії, що уражали до 1,5— 2 млн. чіл.
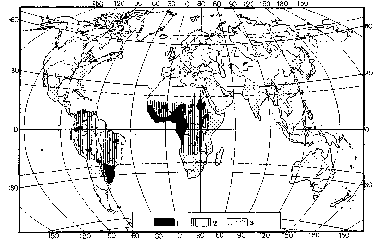
Лихоманка денге зустрічається також у тропічному і субтропічному поясі Північної, Центральної і Південної Америки (Мексика, Гондурас, Коста-Рика, Панама, Бразилія й ін.), у південних штатах США (Техас, Флорида, Каліфорнія, Аризона й ін.), АРІ, Судану, Лівану, Саудівській Аравії, Греції, Південній Іспанії, на о-ві Кіпр (мал. 23).
У місцях постійного поширення лихоманки денге хворіють переважно діти і приїжджі, що залежить від природної імунізації.
2.12 Лихоманка Чикунгунья
Лихоманка Чикунгунья — вірусне захворювання, вперше-описане в 1952—1953 р. у Танганьїці і зустрічається в Східній Африці і Південно-Східній Азії.
Захворювання характеризується появою раптових болів у суглобах і високій температурі, 12-денні інкубаційні періоди. Гострий період хвороби продовжується 6—10 днів, але нездужання може затягуватися до 4 міс. У більшості хворих (80%) спостерігається висипка на тулуб і в області суглобів. Смертельних наслідків у дорослих не описано. Хворіють частіше діти 6—9 років, у даній віковій групі летальність складає 8,2%.
Сезонних змін захворюваності не виявлено, тому що річні коливання температури, вологості й опадів у районі поширення лихоманки Чикунгунья незначні.
Джерелом збудника є хворий у гострій стадії хвороби. Вірус виділений з комарів С. fatigans і Ае. aegypti, виловлених у хатинах, і Ае. africanus, виловлених у лісі.
Вірус передається комарами за допомогою специфічної інокуляції. В організмі комара вірус зберігається до 33 днів.
3. Комарі, переносники збудників бактеріальних хвороб
Комарі пологів Aedes і Culex є також переносниками більшої групи вірусів, розповсюджених у ряді тропічних країн: лихоманки долини Рифт, лихоманки Західного Нілу, О'ньонг-ньонг, Буньямвера й ін.
Крім вірусів, що викликають розглянуті вище захворювання, комарі є також переносниками збудників бактеріальних хвороб і гельмінтозів: туляремії, меліоідозу, філяриатозів.
Малюнок 3.1 – Географічне поширення лихоманки денге: 1 — зони поширення захворювання; 2 — зони поширення переносник -комарів-переносників
3.1 Філяриатози
Філяриатози являють собою групу гельмінтозів, викликуваних гельмінтами підзагону Filariata. Людина для цих гельмінтів є остаточним хазяїном. Проміжні хазяїни — різні види кровосмокчущих комах, переносників цих збудників.
Комарі є переносниками збудників вухерериозу, бругіозу, що характеризуються переважною поразкою лімфатичної системи, що в пізніх стадіях хвороби часто приводить до слоновості різних частин тіла.
3.2 Вухерериоз
Вухерериоз викликається гельмінтом Wuchereria bancrofti, що є двуххозяйним паразитом. Остаточним хазяїном є людина, у якого статевозрілі вухерерії паразитують у лімфатичних судинах і вузлах. Самки філярій відроджують безліч личинок — мікрофілярій (0,12—0,32 мм довжиною), що знаходяться в кровоносних судинах легень, серця, м'язів, в аорті і сонних артеріях, а ввечері і вночі з'являються в периферичній крові. Періодичність надходження мікрофілярій W. bancrofti у периферичну кров підкоряється добовому ритму. Число мікрофілярій у периферичній крові збільшується з 20 ч, досягає піка до 2 ч ночі і падає до мінімального рівня до 10 ч ранку. Стимулами, що обумовлюють періодичність надходження мікрофілярій у кров, є, очевидно, добове коливання парціального тиску кисню в легень і зміну інших фізіологічних функцій хазяїна. Але при зміні людиною способу життя шляхом заміни нічного сну денним мікрофілярій з'являються в судинах шкіри не вночі, а вдень. Крім того, існують штами вухерерії (W. pacifica), мікрофілярій яких зустрічаються в периферичній крові в однаковій кількості і вдень і вночі.
Поява мікрофілярій у периферичних капілярах робить їх .доступними для комарів. У комарі личинки філярій двічі линяють, проробляють складний розвиток і, досягши третьої інвазионної стадії, накопичуються в хоботку. Розвиток мікрофілярій до інвазионній стадії в залежності від температури зовнішнього середовища продовжується від 8 до 35 днів. Оптимальні умови створюються при температурі 21—32° і вологості 70—100%.
Проміжними хазяїнами W. bancrofti і переносниками збудників вухерериозу є різні види комарів пологів Culex Anopheles, Aedes (С. fatigans, С. bitaeniarhynchus, Ае. aegypti, Ае. ро-linesiensis, Ae. vexans, An. gambiae, An. funestus, An. hyrcanus і ін.).
При влученні зараженого комара на людину личинки філярій попадають на шкіру останнього, активно проникають через неї і зі струмом крові заносяться в лімфатичні залози, де досягають полової зрілості. Скупчення філярій можуть викликати закупорку лімфатичних проток, що приводить до застою лімфи, появі набряків і так називаної слонової хвороби. Дорослі вухерерії можуть жити в організмі людини до 17 років; тривалість життя мікрофілярій — близько 70 днів.
Вухерериоз має значне поширення в Африці, Азії, Австралії й Америці. В Азії ураженість вухерериозом і бругиозом спостерігається в 20—80% населення, в Океанії — у 25—58%.
3.3 Бругиоз
Викликається гельмінтом Brugia tnalayi, що також є двуххазяйнним паразитом, але, ,на відміну від W. bancrofti, остаточним хазяїном може бути не тільки людина, але і деякі мавпи (Масаса irus), кішки і собаки. Статевозрілі паразити локалізуються в лімфатичній системі; мікрофілярій з'являються в периферичній крові хворих звичайно вночі, але в деяких районах Далекого Сходу і вдень.
Проміжними хазяїнами і переносниками є комарі пологів Mansonia і Anopheles. Усі переносники філярій антропофильні, однак у тих самих видів ступінь антропофії в різних частинах ареалу різна. Бругиоз має поширення в Південно-Східній Азії, Індії і на Шрі-Ланці. Під час другої світової війни в районі Тихого океану філяріатозами були уражені понад 10000 військовослужбовців.
3.4 Туляремія
Туляремія є гострим інфекційним захворюванням зоонозного характеру, викликуваним Francisella tularensis. Туляремія уражає переважно гризунів, від яких збудник передається людині. Природні вогнища туляремії широко поширені по території Радянського Союзу і ряду закордонних країн.
Збудник туляремії може поширюватися різними шляхами, одним із яких є трансміссивний. Захворювання людей, зв'язані з трансміссивним шляхом передачі спостерігається звичайно в літню пору — з червня до початку вересня. Особливо велике число захворювань відзначається в період достатку комарів і ґедзів, хоча в передачі збудника важливу роль грають також кліщі, блохи і мухи-жигалки. При зараженні через укус кровосмокчущого паразита виникає виразково-бубонна форма туляремії з локалізацією, що відповідає місцеві укусу. На місці укусу з'являється характерна ранка, а в найближчому регіонарному лімфатичному вузлі — бубон. Зараження зв'язане з перебуванням людини в біотопах, характерних для членистоногих і гризунів — резервуарів збудника. У передачі збудника туляремії від тварин людині можуть брати участь практично всі пологи комарів, але найбільше епідеміологічне значення мають представники роду Aedes, що харчуються на гризунах, зокрема водяних пацюках. Вони можуть нападати на хворих пацюків і передавати збудника туляремії здоровим звіркам і заражати людей, що потрапили в даних природне вогнище. Особливо часто збудник туляремії передається комарами Ае. cinereus і Ае. excrucians. Сезон масового літа цих комарів цілком збігається з періодом трансмиссивных спалахів туляремії в середній смузі. Крім зазначених комарів в епідеміології туляремії беруть участь також Ае. vexans, An. hyrcanus, An. maculipennis, С. molestus і інші види.
Механізм передачі збудника туляремії комарами — неспецифічна інокуляція. При кровососании на хворих гризунах на хоботку комарів залишаються збудники туляремії, що можуть зберігатися тут у життєздатному стані до 22—35 днів.
Географічне поширення туляремії обмежене лише північною півкулею. В Америці туляремія виявлена у всіх штатах США, на Алясці, у Канаді, Мексиці і Венесуелі. У Європі вона зустрічається у Франції, Бельгії, Германії і майже у всіх інших країнах; в Азії — у Туреччині і Японії. У СРСР до дійсного часу вогнища туляремії виявлені на заході до границь, на сході до Якутська і Хабаровська, на півночі до Кольськ півострова, на півдні до Криму, Закавказзя, Туркменії і Казахстану включно.
3.5 Меліоідоз чи помилковий сап
Інфекційне захворювання, що не передається від людини до людини, що протікає по типі септикопіемії з утворенням абсцесів у багатьох органах і тканинах (у легень, печінки, селезінці, бруньках, м'язах, суглобах, кістах). Збудник — Pseudomonas pseudomallei. Основні симптоми — швидке підвищення температури до 40—41°, важкі головні і м'язові болі, болі в животі, задишка. Смерть може наступити на 2—15 діб. Летальність — 95—98%.
Резервуаром збудника є пацюки і миші, у яких хвороба може протікати в хронічній формі. Другорядне епідеміологічне значення мають собаки, кішки, коня, корови, свині, морські свинки.
Збудники виділяються в зовнішнє середовище з гнійними відокремлюваним і випорожненнями, що заражають продукти харчування і воду. Зараження людини можливо аліментарним і аерогенним шляхом. Маються вказівки на існування трансміссивній передачі збудника комарами (Ае. aegypti) і блохами (Xenopsylla cheopis).
Вогнища меліоідозу мають поширення в Південно-Східній Азії, Австралії, Африці і Південній Америці.
Висновок
При вивченні зв'язків організмів з фізико-хімічними умовами середовища, що забезпечують обмін речовин, екологія спирається на дані фізіології. При цьому еколога цікавить не життя виду, а процеси, що відбуваються в організмі, пристосувальні особливості видів, їхні життєві форми. Закономірності розміщення видів по території неможливо вивчити без залучення даних біогеографів, геоботаніків, ландшафтоведів, ґрунтознавців, кліматологів. Таким чином, екологія являє собою науку, зв'язану з поруч біологічних і небіологічних наук, що зберігає в той же час свої специфічні задачі й об'єкти дослідження. Значні екологічні особливості характерні для кровосмокчущих комах. Особливості їхня метаморфоза, дивергенція в розвитку, що супроводжується різкою зміною умов життя і гетеротропністю, змушують вивчати екологію окремих фаз розвитку. Личинка, лялечка, доросла комаха мають різну морфологію, різні функції до різні вимоги і зв'язки з зовнішнім середовищем. Важливою задачею екології кровосмокчущих комах і кліщів є з'ясування основних взаємозв'язків з навколишньою природою і людиною.
Предметом еволюційної екології є мікроеволюційні процеси, що протікають у процесі становлення екологічного гомеостазу в осередкових біогеоценозах. Задачі цього розділу екології полягають у виявленні екологічних механізмів еволюційного процесу, шляхів і форм еволюційного становлення індивідуальних, і видових адаптації в біоценотичних системах (Шилов, 1981).Задачами екології комах і кліщів (як і інших тварин) є пізнання формування їхньої морфологічної і фізіологічної особливостей і способу життя в залежності від умов середовища, вивчення впливу середовища на чисельність особ даного виду, на характер розподілу їхній по території і на формування співтовариств організмів, що населяють ту чи іншу територію.
Екологія вивчає ті сторони відносин організмів із середовищем, від яких залежить успішність їхнього розвитку, виживання і розмноження. Вона вивчає не тільки безпосередні взаємини із середовищем, але й історично сформовані на їхній основі специфічні для кожного виду пристосування особ, що забезпечують різнобічні зв'язки із середовищем, їхній розвиток і дозрівання; внутрішньовидові відносини і специфічну для виду структуру його популяції, що визначає його спосіб життя (одиночний, стадний, колоніальний), спосіб використання життєвих ресурсів, розмноження і розселення виду, у результаті чого встановлюються рівень чисельності і характер її динаміки; різні в різних ділянках земної поверхні співтовариства популяцій видів, що зв'язані багатьма взаємними пристосуваннями, що забезпечують біогенний круговорот речовин.
Знання способу життя кровосмокчущих комах і кліщів у різних умовах середовища створює можливість профілактичних заходів, що перешкоджають їх масовому розмноженню. Знання місць випліду комарів, бліх, кліщів і інших членистоногих — переносників збудників хвороб людини і тварин дозволяє цілеспрямовано проводити заходу щодо ліквідації цих трансмісивних хвороб.
Література
Андрианова Н. С. Экология насекомых. — М.: Изд-во Моск. ун-та, 1970. — 158 с.
Артемьев М.М. Комары анофелес — переносчики малярии в СССР//Борьба с ма лярией экологически безопасными методами. — М., 1984. — Ч. 2. — С. 44—60.
Бей-Биенко Г.Я- Общая энтомология. — 3-е изд. — М.: Высшая школа, 1980.— 416 с.
Беклемишев В.Н. Экология малярийного комара (Anopheles macutipennis Wagh.). — М.: Медгиз, 1944.
Бурлаков С.А., Паутов В.Н. Комары и клещи — переносчики возбудителей вирусных и риккетсиозных заболеваний человека. — М.: Медицина, 1975.
Гуцевич А.В., Мончадский А.С., Штакельберг А.А. Комары. Семейство-Си1ісігіае//Фауна СССР: Насекомые двукрылые. — Л.: Наука, 1970. — Т. 3,. вып. 4.
Дажо Р. Основы экологии. — М.: Прогресс, 1975. — 416 с.
Дре Ф. Экология. — М.: Атомиздат, 1976. — 165 с.
Коренберг Э.И., Юркова Е.В. Проблема прогнозирования эпидемического про явления природных очагов болезней человека//Мед. паразитол. _и паразит, болезни. — 1983. — С. 3—10.
Лебедева Н.Н., Фильчагов А.В. Новые данные по экологии личинок lx.od.es-ricinus//Пятое Всесоюзное акарологическое совещание. — Фрунзе, 1985. — С. 186—187.
Мазохин-Поршняков Г.А. Зрение у насекомых. — М.: Наука, 1965.
Никольский Г.В. О содержании, теоретических основах и основных задачах эко логии животных//3оол. журн. — 1955. — Т. 31, вып. 1.
О дум Ю. Основы экологии. — М.: Мир, 1986. — Т. 1. — 328 с. — Т. 2, — 376 с.
Олсуфьев Н.Г., Дунаева Т. Н. Природная очаговость, эпидемиология и про филактика туляремии. — М.: Медицина, 1970.
Руководство по медицинской энтомологии. —• М.: Медицина, 1974.Руководство по тропическим болезням/Под ред. проф. А.Я. Лысенко. — М.: Меди цина, 1983.
Риклефс Р. Основы общей экологии. — М.: Мир, 1979. — 424 с.
Росс Г., Росс Ч., Росс Д. Энтомология. — М.: Мир, 1985. — 572 с.
Руководство по медицинской энтомологии/Под ред. В. П. Дербеневой-Уховой. — М.: Медицина, 1974. — 360 с.
Шилов И.А. Биосфера, уровни организации жизни и проблемы экологии//Экология. — 1981. — № 1. — С. 5—11.
Шовен Р. Мир насекомых. — М.: Мир, 1970. — 840 с.
Яхонтов В.В. Экология насекомых. — М.: Высшая школа, 1969. — 488 с.

Нравится материал? Поддержи автора!
Ещё документы из категории экология:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ