Генетическая история человечества
 Генетическая история человечества
Генетическая история человечества
Доктор биологических наук Лев Животовский, доктор биологических наук Эльза Хуснутдинова
Мы не единожды предлагали нашим читателям статьи, посвящённые проблемам эволюции и развития Homo sapiens. Сегодня мы обратимся к новому аспекту вопроса — генетической истории человечества и его родственников. Каковы пути эволюции с точки зрения генетики? Насколько отличаются ДНК человека и его человекообразных родственников? Кто наши предки и кем нам приходится неандерталец? Ответы на эти и другие вопросы попытались дать ведущие российские биологи.
Эволюция и филогенетическое древо
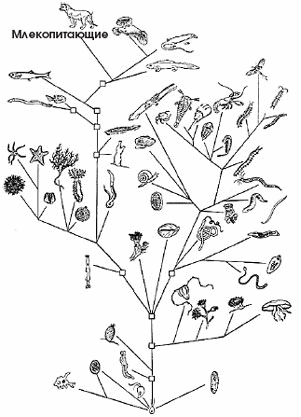
Эволюцию живых существ можно представить в виде величественного раскидистого дерева, корни которого скрыты от нас в глубине веков. Говоря о биологической эволюции, мы рассматриваем лишь надземную часть генетического древа, которая, согласно всем современным научным данным, развивалась постепенно — как растёт любое дерево. Сначала идёт ствол, потом — большие ветви, от них — более мелкие ветки, затем веточки и так далее (см. рис. 1).
Рис. 1. Филогенетическое древо животных (по: Матвеев, 1935).
Нам не дано видеть ветвей эволюционного древа — их составляют давным-давно исчезнувшие виды, жившие десятки и сотни миллионов лет назад. Многие из них не оставили даже следа в палеонтологической летописи, а о других нам рассказывают ископаемые находки. Реально мы наблюдаем только густую крону, где листочки — ныне существующие виды, одним из которых является биологический вид Homo sapiens, рода Человек (Homo) семейства Люди (Hominidae) отряда Приматы (Primates) класса Млекопитающие (Mammalia).
Научным методом изучения эволюции является выявление родственных (филогенетических) связей между различными организмами. В основу исследований, начиная с Карла Линнея (XVIII век), был положен принцип схожести (или несхожести) ныне существующих форм живых существ по фенотипу. Организмы, близкие по морфологическим признакам, физиологическим характеристикам, особенностям развития и др., группируются в одни веточки, другие, отличающиеся от первых, но схожие между собой, — в другие, которые затем образуют всё более крупные ветви. На рис. 1 представлено филогенетическое древо, отражающее фенотипическое сходство разных видов.
Эволюционная теория Чарлза Дарвина и филогенетические методы Эрнста Геккеля позволили современной биологии рассматривать филогенетические древа как эволюционные схемы. Согласно этим представлениям, внутри каждого вида может образоваться разновидность, наделённая новыми особенностями, позволяющими по-иному приспосабливаться к среде обитания. Так на эволюционном древе обозначается рост новой веточки. Если вновь приобретённые качества наследуются, то генетические различия усиливаются в следующих поколениях за счёт отбора свойств, обеспечивающих выживание генотипов, и накопления новых мутаций. Разновидность лучше приспосабливается к существующим условиям, фенотипически удаляясь от родительского вида Так появившаяся веточка растёт, обособляясь в новый вид.
Эволюция и ДНК
Как генетика помогла в интерпретации филогенетического древа и в понимании процесса эволюции? Дело в том, что биологическая эволюция во многом связана с изменением ДНК, представляющей собой последовательность четырех химических соединений — нуклеотидов A, T, C, G (аденин, тимин, цитозин, гуанин). Все ДНК организма называется геномом. Определённые участки ДНК, гены, кодируют белки; имеются также некодирующие участки генома. Это и есть генетический текст, определяющий как видовые признаки, общие для всего вида, так и уникальные особенности, отличающие данную особь от других представителей того же вида. ДНК любого организма подвержена мутациям, часть которых не изменяют числа нуклеотидов на данном участке ДНК, а меняют их местами. Но возможны и более сложные процессы: выпадения, вставки, удвоения нуклеотидов и перемещения фрагментов ДНК из одной части генома в другую; не исключён даже перенос ДНК между разными видами.
Мутация — редкое событие. Вероятность того, что данный нуклеотид в ДНК потомка будет изменён по сравнению с родительским, равна примерно 10–9. Однако для громадных промежутков времени, на протяжении которых разыгрывается эволюционный процесс, для всего генома, состоящего из огромного числа нуклеотидов (у человека их 3 млрд.), это ощутимая величина. Особи с вредными для организма нарушениями не выживают или не участвуют в размножении, и мутации далее не передаются. Полезные же изменения могут наследоваться потомками: так из поколения в поколение генетическая информация преобразуется — в этом и заключается генетическая суть процесса эволюции.
На филогенетическом древе внешне сходные друг с другом виды сгруппированы на одной ветви. Изучение ДНК ныне живущих видов позволило сопоставить близость особей разных видов на уровне эволюционных изменений, вызванных мутациями. Современная молекулярная биология позволяет сравнить соответствующие фрагменты ДНК (скажем, определённого гена) у разных видов и подсчитать число различий между ними. Филогенетические древа, построенные как по ДНК, так и по морфофизиологическим признакам, имеют очевидные соответствия: виды, далёкие друг от друга на морфофизиологическом древе, так же далеки на ДНК-филогенетическом. Таким образом, генетика доказала, что классическое филогенетическое древо отражает направление эволюционных преобразований. Более того, она показала, какие именно изменения геномов сопровождают эволюцию каждой из таксономических групп.
Человек и другие виды
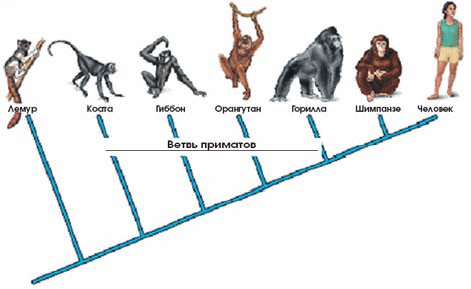
Рис. 2. Чем больше времени прошло с момента разделения видов, тем больше накопилось различий в их ДНК.
Сравним человека, скажем, с бабочкой. Очевидно, что мы очень отличаемся друг от друга по внешнему облику и по составу ДНК и находимся на далёких ветвях филогенетического древа. Перейдём теперь к млекопитающим. Если сопоставить человека, скажем, с кошкой или собакой, к которым мы гораздо ближе, чем к бабочкам, то окажется, что и по ДНК человек более схож с ними. Если отправиться по ветви млекопитающих дальше, к приматам, то по мере приближения к человеку родственные черты с человекообразными — орангутаном, гориллой и шимпанзе — становятся очевидными (рис. 2). Больше всего человек походит на шимпанзе. Если сопоставить ДНК, окажется, что они очень близки. Генетика позволила количественно оценить сходство: человек и шимпанзе отличаются друг от друга всего одним-двумя нуклеотидами из каждых ста. То есть генетическое тождество составляет чуть ли не 99%.
Люди генетически близки друг к другу
Перейдём теперь собственно к человеку. Сопоставим представителей таких далёких народов, как аборигены Океании и европейцы, или сравним лица всем известных людей. Очевидно, что они разные, но насколько?
Сумел бы прилетевший на Землю инопланетянин отличить нас друг от друга, или мы все показались бы ему на одно лицо? Ведь случайному посетителю зоопарка все мартышки кажутся одинаковыми, а человеку, работающими с ними, — абсолютно разными. Конечно, неандерталец имеет мало общего с современным человеком, но стоит „одеть“ его в костюм и шляпу, он становится одним из нас. Автор этой статьи на лекции по теории эволюции для школьников спросил, кто изображён на рисунке. И тут же один из ребят воскликнул в радостной догадке: „Так это же вы!“
Если сравнить ДНК разных людей, то выяснится, что они отличаются друг от друга лишь на 0,1%, то есть только каждый тысячный нуклеотид у нас разный, а остальные 99,9% совпадают. Более того, если сопоставить всё разнообразие ДНК представителей самых разных рас и народов, то окажется, что люди отличаются гораздо меньше, чем шимпанзе в одном стаде. Так что гипотетический инопланетянин сначала научится отличать друг от друга шимпанзе, а лишь затем — людей.
Много это или мало — 99,9% сходства и 0,1% различий. Проведём простые подсчёты. ДНК человека содержит около 3 млрд. пар нуклеотидов, примерно три миллиона из них у каждого из нас разнятся. Этого достаточно, чтобы утверждать, что не существует людей, генетически тождественных друг другу. Даже ДНК близнецов могут отличаться вследствие мутаций. Правда, большинство различий приходится на молчащие участки ДНК, и потому основные гены у нас во многом идентичны. Рассмотрим, например, молекулу гемоглобина, играющую ключевую роль в транспортировке кислорода из лёгких в клетки организма. Состав данной молекулы у всех абсолютно одинаков. Конечно, единичные отклонения возможны, но все они сопровождаются серьёзной патологией, поскольку мутация хотя бы одной аминокислоты в сложной молекуле гемоглобина изменяет её конфигурацию, резко нарушая способность удерживать кислород и снабжать им организм. Подобным же образом у всех людей совпадает множество других белков и кодирующих их генов.
Часть нуклеотидов, отличающих нас друг от друга, наделяет людей рядом признаков, определяющих группы крови, телосложение, цвет кожи, поведение и др. и позволяющих приспосабливаться к меняющимся условиям жизни. Однако большинство различий не связано напрямую с приспособительными функциями организма, их эволюционный ход определяется скоростью мутационного процесса, что позволяет проследить пути эволюции человека и его расселения по земному шару.
Предки человека
Так почему же особи шимпанзе больше отличаются, чем люди? Почему мы так похожи друг на друга генетически? Откуда мы и кто наш предок? Последний вопрос до сих пор остаётся спорным, хотя археологические находки последних десятилетий и исследования ДНК приблизили нас к пониманию этого. Шимпанзе имеет длительную историю развития, в ходе которого выработалось значительное генетическое разнообразие. Эволюционная же история человека слишком коротка для накопления серьёзных различий. Обратимся теперь к деталям нашего прошлого.
5–7 млн. лет назад от общего предка человека и шимпанзе отделилась ветвь, давшая начало древним обезьяно-людям, переходящим к прямохождению. Обитали они в Южной Африке, где их останки и были найдены; отсюда и их название — австралопитеки (т. е. обезьяны из Южного полушария), род Australopithecus. Одна из ветвей дала начало роду Человек — Homo. До середины 90-х г. древнейшим известным предком современного человека считался афарский австралопитек, A. afarensis, живший 3–4 млн. лет назад. К этому виду относится знаменитая Люси, найденная в 1974 г. в Эфиопии. Ряд палеоантропологов полагают, что древнейшим прародителем современного человека являлся другой вид австралопитека — рамидус, A. ramidus, чьи останки, насчитывающие 4,5 млн. лет, также были обнаружены в Эфиопии.
Первый известный нам представитель рода Человек появился более 2 млн. лет назад. Это был Homo habilis — Человек умелый, живший в Восточной и Южной Африке. Потом его сменил Homo erectus — Человек прямоходящий, следы которого были найдены в Африке и Евразии — им 1,7 млн. лет. К этому виду принадлежит азиатский синантроп, или пекинский человек, питекантроп, или яванский человек, обнаруженный в Океании, и древний „европеец“ — гейдельбергский человек. Самым поздним находкам — 250 тыс. лет. Антропологи сходятся во мнении, что Homo erectus распространился далеко за пределы Африки путём миграций.
Первые представители Homo sapiens (т.н. архаичные люди) являющиеся потомками Homo erectus, появились более 500 тысяч лет назад и отличались по сложению от человека современного типа. Их кости были обнаружены и в Африке, и в Европе, и в Азии. И поэтому возникает вопрос: а где же возник современный человек? В настоящее время существуют две основные гипотезы: полифилетическая (или мультирегиональная) и монофилетическая.
Рис. 3. Мультирегиональная гипотеза. Человек современного анатомического типа возник независимо на разных континентах из разных ветвей Homo erectus. Единообразие людей поддерживалось системой миграций (по: Stringer, 1989).
Рис. 4. Монофилетическая гипотеза (гипотеза африканского происхождения человека). Человек современного типа возник в Африке и распространился оттуда на все континенты, вытесняя других представителей рода Homo (по: Stringer, 1989).
Согласно мультирегиональной гипотезе (рис. 3), люди разных рас имеют различное происхождение: каждая возникла на своём континенте от отдельного представителя вида Homo erectus. При этом предковые расы составляли единую популяционную систему и, мигрируя, обменивались генами. Монофилетическая же гипотеза утверждает, что все люди имеют общее происхождение, и Homo sapiens, как и Homo erectus, возник в Африке, а затем расселился оттуда по всем континентам .
Сторонники обеих гипотез не сомневаются в происхождении человека от Homo erectus, но расходятся в вопросе о времени его возникновения. Согласно мультирегиональной гипотезе, возраст современного человека более миллиона лет, а по монофилетической версии — около 500 тыс. Однако ни антропологи, ни археологи не смогли поставить точку в этом споре. Коренной перелом в решении проблемы возникновения человека, его расселения по континентам и возникновения рас внесли исследования ДНК.
ДНК и восстановление истории человечества
Историю популяций человека можно проследить, сравнивая представителей разных рас и народов по тем фрагментам ДНК, по которым различия между индивидами гораздо более выражены, чем в среднем по геному. Такие фрагменты называют ДНК-маркёрами. Зная скорость, с которой происходят мутации, и определяя количественную меру различий между геномом людей из разных популяций, вычисляют время их отделения от общей предковой линии. Продвигаясь назад от генома современного человека, можно построить филогенетическое древо Homo sapiens. Оно позволяет делать определённые выводы о генетической истории человечества.
Рис. 5. Фрагмент ДНК. Разные индивиды могут отличаться друг от друга по составу нуклеотидов (при постоянной длине фрагмента) или по длине фрагмента (за счёт вставки или выпадения нуклеотидов).
Для исследования используют разные типы ДНК-маркёров. Следует отметить ДНК митохондрий (мтДНК) и Y-хромосомы (Y-ДНК), поскольку они позволяют проследить генетическую историю человечества отдельно по женской и мужской линиям. Первая передаётся только по материнской линии, так как митохондрии находятся в цитоплазме клетки, а цитоплазма потомка (зиготы) определяется цитоплазмой яйцеклетки. Если два человека имеют общего предка женского пола, то различия между их мтДНК покажут, сколько поколений отделяет их от жившей столетия или тысячелетия назад общей прабабушки. Изучение Y-ДНК позволяет проследить эволюционные траектории по отцовской линии, поскольку Y-хромосома передается только по мужской линии. Изучение остальной, аутосомной части генома, которая сосредоточена в хромосомах, наследуется по обеим линиям и в которой представлена большая часть генома человека, также чрезвычайно важно, ибо даёт маркёры для изучения комбинативной изменчивости, одновременно привносимой и с отцовской, и с материнской сторон.
Первое исследование такого рода было проведено с использованием митохондриальной ДНК. Сравнивая данные аборигенов всех континентов, учёные обнаружили, что разнообразие мтДНК выше всего в Южной Африке. Более того, там были обнаружены такие типы, которые нигде больше не встречались. Они оценивались по нуклеотидному составу как наиболее древние. Митохондриальные ДНК населения других континентов были менее разнообразны, их сравнение с мтДНК аборигенов Южной Африки показало, что они возникли как мутационные изменения африканских типов в ходе распространения человечества за пределы Африки.
Изучение географического распределения типов ДНК-маркёров и анализ их мутационных взаимосвязей позволили доказать справедливость монофилетической гипотезы (рис. 4). Более того, их изучение помогает реконструировать миграционные события не только далёких тысячелетий, но и последних веков. Например, эпоха великих географических открытий ХIV-XVI вв. способствовала развитию контактов с неведомыми ранее жителями отдалённых земель. На кораблях были только мужчины, и сейчас генетические исследования аборигенных народов Африки, Океании и Америки показывают присутствие в их ДНК значительной доли типов Y-хромосомы, характерных для европейцев.
«Митохондриальная Ева» и «Y-хромосомный Адам»
По степени разнообразия ДНК-маркёров можно установить, от какого предкового типа и когда они произошли. Более того, поскольку различные типы мтДНК не рекомбинируют, т. е. не обмениваются между собой фрагментами в процессе образования половых клеток, то вероятностный анализ показывает, что все ныне существующие типы мтДНК сводятся в далёком прошлом к одному-единственному прототипу. Почему это происходит? Допустим, существовала небольшая прапопуляция. Если одни женщины в ней имели больше детей, чем другие, то в следующем поколении именно их мтДНК будет встречаться чаще. Их дочери и внучки тоже имели детей, унаследовавшх мтДНК своих предков. При этом случайным образом возникают мутации. Так развивается стохастический процесс в передаче типов мтДНК, в результате чего из поколения в поколение одни типы мтДНК увеличивают свою представительность в популяции, другие — уменьшают.
Согласно теории стохастических процессов, когда-нибудь в популяции останутся потомки только одной мтДНК — от некой единственной женщины, образно именуемой „митохондриальной Евой“. Но и её соплеменницы внесли свой вклад в генофонд отдалённых потомков, поскольку наш геном содержит не только митохондриальную ДНК. Длина мтДНК лишь около 16,5 тыс. нуклеотидов, что ничтожно мало по сравнению с 3 млрд. нуклеотидов ДНК хромосом, основная часть которых представлена 22 аутосомами и X-хромосомой, где комбинируется наследственность по обеим линиям, женской и мужской. Из-за многочисленных мутаций мтДНК ныне живущих людей отличается от данных „Евы“, возраст которой (т. е. точки соединения всех типов мтДНК в прошлом) примерно 200 тысяч лет. Аналогичная ситуация и с Y-хромосомой. Большая часть её не рекомбинирует, и изменчивость ДНК в ней подчиняется тем же стохастическим процессам, что и мтДНК. При этом говорят о „Y-хромосомном Адаме“. Он оказался гораздо моложе „мтЕвы“. Объяснение тому — меньшая, т. н. эффективная численность мужчин на протяжении истории человечества из-за их большей смертности и нередкого отстранения от процесса размножения.
Неандертальский человек — наш прародитель или двоюродный дядя? Дополнительные сведения о нашей родословной были получены при сравнении митохондриальных ДНК современного человека и неандертальца. Более ста лет, с тех пор как в Германии впервые были найдены останки нашего древнего родственника, шли дискуссии о том, кем он нам приходится. Исходя из особенностей строения скелета неандертальца и его географической распространённости, одни учёные считали его нашим прародителем, т. е. линией Homo sapiens, развитие которой привело к человеку современного анатомического типа. Другие детали позволяли считать его независимой эволюционной ветвью, подвидом Homo sapiens, имеющим общего с нами предка, т. е как бы двоюродным дядей. Эти два подвида получили зоологическое название Homo sapiens neanderthalensis и Homo sapiens sapiens.
Учёным удалось прочитать часть вариабельного (контрольного) участка митохондриальной ДНК двух неандертальцев. Первый был найден в Фельдховеровской пещере в Германии, чуть позже был прочитан генетический текст мтДНК неандертальца-ребёнка, обнаруженного на Северном Кавказе в Мезмайской пещере. При сравнении наиболее изменчивых фрагментов мтДНК современного человека и неандертальца были найдены существенные различия: они отличались друг от друга в среднем по 27 нуклеотидным позициям из 370 исследованных. Если сравнить митохондриальные ДНК двух современных людей, то средняя разница будет равна лишь 8 нуклеотидам. Эти расчёты показывают, что наш общий с неандертальцем предок жил примерно 500–700 тыс. лет назад.
Анализ ДНК позволил заключить, что обмен генами между человеком и неандертальцем не происходил или был ничтожно мал. Вероятнее всего, это совершенно отдельные, параллельные эволюционные ветви, произошедшие от общего предка. Хотя для окончательного вывода нужны дополнительные исследования ДНК. Так что неандерталец нам, похоже, эволюционный „дядя“. Приблизительно 300–400 тыс. лет назад произошло окончательное разделение двух ветвей. Неандертальцы первыми расселились по Европе и Азии, затем туда пришли люди современного типа (т.н. кроманьонский человек), и они довольно долго сосуществовали на одной территории. Но около 30 тыс. лет назад неандертальский человек исчез, никаких его следов в более поздних археологических слоях найдено не было. Возможно, он не выдержал конкуренции и был вытеснен и истреблён своим более умным и хитрым родственником, а может быть, существовали и иные причины гибели неандертальцев.
Эволюция популяций и рас
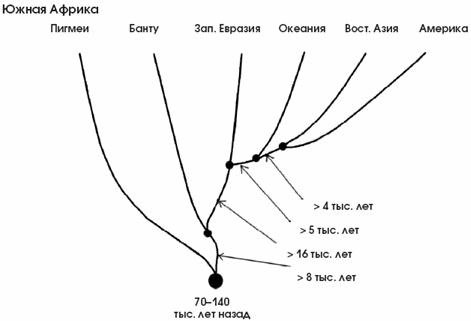
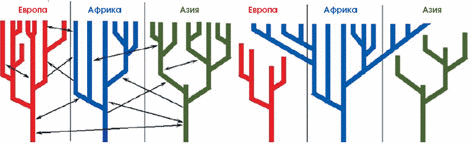
Рис. 6. Эволюционное древо популяций человека, по данным о ДНК-маркёрах. Видно, что первично развитие и отделение друг от друга популяций началось в Африке в среднем около 100 тыс. лет назад. Затем одна ветвь вышла из Африки и стала делиться на континентальные ветви. Стрелками указано минимальное время, прошедшее между отделением эволюционных ветвей. Следует иметь в виду, что отделение ветви не означает ещё физического присутствия популяций в этом регионе. Например, ветвь, ведущая от азиатских популяций к американским индейцам, показывает, когда эта ветвь отделилась, но нужно было ещё время, за которое отделившиеся группы достигнут Берингии и перейдут пролив (по: Zhivotovsky et al., 2003).
Изучая разнообразие ДНК современных народов, можно оценить численность той прапопуляции, от которой, согласно гипотезе африканского происхождения, произошло всё человечество. Она была невелика — порядка нескольких тысяч. Сопоставляя ДНК-маркёры аборигенов Южной Африки, можно сказать, что примерно 70–150 тыс. лет назад началась интенсивная дифференциация и сложные демографические процессы, сопровождающиеся возникновением разнообразных популяций в пределах Африки. Затем, 50–100 тыс. лет назад, волны переселенцев стали выплёскиваться за пределы Африки и растекаться по другим континентам, что отразилось на своеобразной структуре ДНК-древа (рис. 6).
Исследуя современное население Европы, Азии, Океании, Северной и Южной Америки и зная особенности и скорость мутирования в изучаемых ДНК-маркёрах, можно с определённой степенью точности проследить пути и время миграций людей из Африки. Удивительно то, что генетические данные подтверждаются археологическими находками. Например, структура ДНК свидетельствует о том, что человек появился в Австралии и Новой Гвинее 50–60 тыс. лет назад. Анализ состава химических элементов артефактов указывает на тот же период. В Центральную и Юго-Восточную Азию люди пришли примерно 70 тыс. лет назад, заселение Европы произошло позже, 35–40 тыс. лет назад. Время освоения Америки до сих пор не определено, известно лишь, что люди появились там гораздо позже, чем на других континентах, — от 15 до 35 тыс. лет назад.
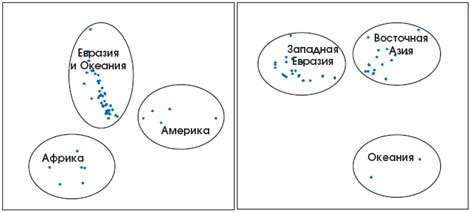
Рис. 7. Подразделение этнических группировок по географическим районам, осуществлённое по ДНК-маркёрам. Каждая точка представляет собой выборку индивидов определённой этнической группы из данного географического региона, охарактеризованным по четыремстам аутосомным ДНК-маркёрам. Левый и правый рисунки представляют собой разные проекции многомерного пространства различий между выборками. В каждом из регионов прослеживается группировка этнических групп общего происхождения, однако различия между ними уже не столь яркие, как между регионами (по: Zhivotovsky et al., 2003).
Как возникли современные расы человека и отличаются ли они друг от друга по ДНК? В течение десятков тысяч лет шли процессы миграций и адаптации человека к местным условиям. Допустим, группа людей пришла в Юго-Восточную Азию и осела там на много поколений. Потом часть мигрировала дальше, образуя новую локальную популяцию, которая, однако, имеет общую историю и общих предков с родительской группой, а потому их ДНК более сходны между собой, чем с жителями других континентов. Действительно, население разных материков эволюционно гораздо дальше от общей предковой группы, чем соседние популяции, близкие по родственным связям и демографической истории. За то время, что прошло со времени отделения от общих прародителей, их ДНК стали отличаться друг от друга за счёт накапливающихся в чреде поколений мутаций. Генетические различия между людьми с разных материков мы называем сегодня расовыми признаками. Изучая десятки и сотни ДНК-маркёров, можно почти стопроцентно идентифицировать расу (рис. 7). Чтобы достоверно определить этническую принадлежность индивида в пределах расы и крупного географического региона, потребуются тысячи ДНК-маркёров. А в зонах контакта разных рас и этнических групп это сделать практически невозможно из-за смешения генофондов.
Но генетически мы все вышли из одного гнезда, причём сравнительно недавно в масштабах эволюции (рис. 6).
Дальнейшее развитие рас шло независимо друг от друга: люди адаптировались к климато-географическим условиям, типу питания и ландшафта, складывались язык и культура. Но на формирование народов влияли не только процессы разделения популяций. Новые этносы могли образоваться при смешении групп разной расовой и языковой принадлежности. При этом возникала генетически разнородная этническая общность с единым типом культуры и общим языком. Поэтому сейчас всё большую актуальность приобретают исследования, связанные с изучением генофонда, т. е. всего разнообразия ДНК в популяциях, генетической истории населения отдельных регионов, расово-этнических групп, родословной современных этносов.
Контакт этносов
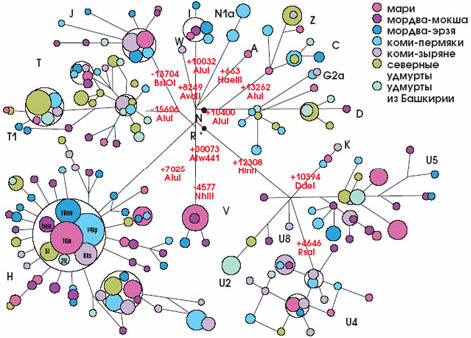
Особый интерес с этой точки зрения представляет Волго-Уральский регион — в силу особенностей этнической истории населяющих его народов. Здесь встретились две волны расселения, две расы: европеоидная и монголоидная. Следы этого события хранит ДНК проживающих здесь народов (рис. 8). Исследования митохондриальной ДНК и Y-хромосомы позволили рассчитать время формирования народов, заселивших впоследствии данный регион. Это произошло примерно 40–50 тыс. лет назад, что соответствует времени появления современного человека на европейском континенте в эпоху верхнего палеолита.
Рис. 8. Сеть мтДНК народов Волго-Уральского региона. Цветом обозначены различные этнические группы, тёмными буквами — разные типы мтДНК, размер кружка — число индивидов с данным типом мтДНК, красными цифрами — мутации. Видно значительное перекрывание изменчивости: в различных этнических группах встречаются одинаковые типы мтДНК, в т. ч. европеоидные, напр. Н., и монголоидные, напр. U. (из: Бермишева и др., 2002).
Сравнительный анализ мтДНК 18 народностей Евразии, относящихся к тюркской ветви алтайской языковой семьи, позволил установить западно-восточный градиент увеличения частоты азиатских типов мтДНК на пространстве 8 тыс. км: от 1% у гагаузов из Молдавии до 95–99% у якутов и долган. Соответственно, европеоидные черты наиболее свойственны жителям Западной Евразии, а наименее — населению Восточной Сибири. Народы Волго-Уральского региона, а также узбеки и казахи, т. е. те, кто живёт на границе между Европой и Азией, занимают промежуточное положение. Изучение аутосомных ДНК-маркёров показало наличие в генофонде народов Волго-Уральского региона значительной доли европеоидных черт — от 50 до 90%. Таким образом, оказавшись на границе между Европой и Азией, эти народы сохранили следы смешения двух рас, пришедших одна — с Востока, а другая — с Запада. Кроме того, оказалось, что сходство языков играет меньшую роль, чем географическая близость популяций. Если, например, русские из Рязанской и Курской областей имеют только 2–3% монголоидных типов мтДНК, то русские, проживающие на границе Европы и Азии, имеют их уже 10–12%. Это объясняется их смешением с тюркоязычными народами на территории Волго-Уральского региона.
Таким образом, ДНК какого-либо индивида не даёт возможности определить его национальность, но позволяет выяснить, какого типа у него мтДНК или Y-хромосома: скажем, монголоидной или европеоидной линии. У русских Волго-Уральского региона есть ДНК-маркёры, которые характерны и для марийцев, и для мордвы, и для чувашей, и для башкир, и для татар, и для удмуртов; соответственно, марийцы имеют ДНК-маркёры, встречающиеся у других этнических групп Волго-Уральского региона, и т. д. Эти результаты показывают глубокую генетическую общность давно живущих рядом народов, хотя говорят они на непохожих языках, веруют в разных богов и отличаются культурными традициями.
Все люди — генетические кузены и кузины
Мы все генетически чрезвычайно схожи. Нас отличает друг от друга ряд признаков, которым мы склонны придавать чересчур большое значение (рост, цвет кожи, форма головы и др.), но как они ничтожны в сравнении с нашим почти стопроцентным (99,9%) генетическим сходством! Мы сформировались под влиянием не только генов, но и окружающих нас людей и явлений, причём в становлении личности среда играет неизмеримо большую роль, чем наследственные особенности. Мы все — одна большая генетическая семья, живущая на общей планете. И все распри между людьми возникают на бытовой почве: из-за несоблюдения элементарных норм общежития народов, неуважения к ценностям, особенностям и кажущимся странностям друг друга.
Но представьте себя на необитаемом острове, куда случай забросил ещё одного человека — совсем другой расы, другого вероисповедания, со своим языком и привычками, — 99,9% генетического сходства вас тут же объединят.

Нравится материал? Поддержи автора!
Ещё документы из категории география:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ