Журнал высшей нервной деятельности том 40 1990 вып. 3



 ЖУРНАЛ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
ЖУРНАЛ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
ТОМ 40 1990 ВЫП. 3
УДК 612.821.6+612.822.3+615.78 © 1990 г.
АЛЕКСАНДРОВ Ю. И., ГРИНЧЕНКО Ю. В., СВЕТЛАЕВ И. А.
ВЛИЯНИЕ ОСТРОГО ВВЕДЕНИЯ ЭТАНОЛА НА РЕАЛИЗАЦИЮ ПОВЕДЕНИЯ И ЕГО НЕЙРОННОЕ ОБЕСПЕЧЕНИЕ
В экспериментах на кроликах, обученных инструментальному пищедобывательному поведению, выясняли, какие изменения активности нейронов лимбической области коры соответствуют нарушению этого поведения (увеличение времени реализации и числа ошибок), вызванному внутрибрюшинным введением 12%-ного раствора этанола в дозе 1 г/кг. По сравнению с контролем (введение изотонического раствора) число активных клеток, выделяемых в микроэлектродном треке, уменьшилось на 1/3; паттерн поведен ческой специализации нейронов, вовлекающихся в обеспечение нарушенного поведения, изменился. Содержание нейронов наиболее новых систем, сформированных при обу чении животных инструментальному поведению, уменьшилось с 27 до 11%, а нейронов, обеспечивающих реализацию систем, сформированных на предыдущих этапах инди видуального развития, увеличилось с 18 до 36%.
Влияние острого введения этанола на разные формы поведения жи вотных и человека изучено в большом числе работ (см. обзор [10]). Имеется также много данных о действии этанола на медленную и им пульсную активность нервной клетки, полученных в аналитических экс периментах на разного рода препаратах, культуре ткани (см. обзоры [16, 25]). Казалось бы, в литературе имеются необходимые компоненты для развития представления о нейронных основах действия этанола на поведение. Однако ситуация осложняется тем, что острое влияние эта нола на активность нейронов не может быть названо ни «общевозбуж дающим», ни «общетормозным» [21]. Разнонаправленность действия этанола обнаружена не только для нейронов разных структур, но и для нейронов внутри одной структуры [15, 18, 25]. Кроме того, действие этанола на активность нейронов зависит от целого ряда факторов: до зы, концентрации в крови и ликворе, способа введения, вида наркоза [15, 18, 20, 24]. Поэтому для сопоставления двух групп данных — о дей ствии этанола на поведение и на активность нейронов, надо иметь до статочно информации, чтобы учесть названные факторы, быть уверен ным в том, что действия этанола на активность данного нейрона в ус ловиях аналитического эксперимента и в конкретной форме поведения совпадают, а также знать, какова роль соответствующих групп нейро нов в обеспечении данной формы поведения. Возможность соблюдения всех этих условий по меньшей мере сомнительна. Реально даже лучшие попытки подобных сопоставлений [12] основываются лишь на самых общих представлениях о механизмах поведения, в понимании которых существуют значительные расхождения позиций у разных авторов.
С учетом этих, а также других препятствий, возникающих при сопо ставлении данных поведенческих и нейрофизиологических эксперимен тов [14], становится очевидным, что оптимальным путем выяснения нейронных основ действия этанола является постановка эксперимента, в котором проводится сопоставление вызванных этанолом изменений активности нейронов с его поведенческими эффектами. Задача настоя щего исследования состояла в том, чтобы выяснить, какие изменения организации нейронной активности соответствуют вызванному острым введением этанола нарушению поведения. В экспериментах на кроли ках, которые были объектом наших исследований, показано, что наи более чувствительны к действию этанола палео- и неокортикальные об разования, в том числе структуры лимбической системы [18]. Считает ся, что реорганизация активности лимбических структур является су щественным звеном в механизмах формирования потребности в алкого ле [8]. Мы исследовали активность нейронов области 29d (корковый
456
отдел лимбической системы), которая выделяется с морфологической точки зрения богатством связей с другими областями коры и особой стратегической позицией структуры, связывающей неокортикальные об ласти и гиппокамп [23]. В лимбической коре обнаруживается макси мальное по сравнению с другими областями коры количество термина-леи, содержащих дофамин [13], нарушение обмена которого играет, по-видимому, ключевую роль в патогенезе алкоголизма [5].
МЕТОДИКА
Эксперименты проведены на трех взрослых кроликах-самцах, реа лизующих подробно изученное ранее инструментальное пищедобыва-тельное поведение [2, 3]. Экспериментальная камера была оборудова на двумя педалями и двумя автоматически подающимися кормушками. Педали располагались у задней стенки в правом и левом углах. У пе редней стенки в правом и левом углах находились кормушки. При на жатии на левую педаль подавалась левая кормушка, на правую — пра вая (подробнее см. [2]).
Животных обучали сначала захвату пищи из правой кормушки, за тем нажатию на правую педаль. В той же последовательности обучали поведению у левой стенки.
После того как поведение кроликов стабилизировалось в обоих пи-щедобывательных циклах (у правой и левой стенок): вслед за захватом порции пищи из кормушки животное направлялось к педали, нажима ло на нее, подходило к кормушке, захватывало пищу и т. д.,— переходи ли к экспериментам с введением этанола (ЭЭ). Этанол (12% в изото ническом растворе) вводили внутрибрюшинно в дозе 1 г/кг и затем че рез каждые 1,5—2,0 ч по 0,3—0,5 г/кг в зависимости от индивидуаль ных особенностей метаболизма и режима потребления пищи животными. Концентрация алкоголя в крови определялась методом газовой хрома тографии [4].
В контрольных экспериментах (КЭ) вводили эквивалентные количе ства изотонического раствора. Между прекращением предыдущего и началом следующего ЭЭ проходило минимум 65—70 ч.
Способы регистрации, обработки импульсной активности, а также критерии выделения активаций и классификации нейронов были описа ны ранее [2, 3]. Отметим основное. Активность нейронов и т. masseter, служебные отметки регистрировались на магнитную ленту магнитогра фа НО-46. Параллельно записывали поведение животного и импульсную активность с помощью видеомагнитофона «Электроника-509».
У каждого животного сопоставляли время реализации циклов пове дения (критерий t) и количество ошибок совершения поведения («Хи-квадрат») в КЭ и ЭЭ. Различия считались достоверными при p<0,05. В тех же экспериментах регистрировали активность нейронов области 29d лимбической коры (латеральный отдел на уровне Р 10,0 в соответ ствии с [23]). Достоверность изменений числа нейронов, принадлежа щих к разным классификационным группам, оценивали по критерию «Хи-квадрат» и по точному критерию Фишера.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Концентрация алкоголя в крови достигала максимума 0,9±0,24 г/л через 15—20 мин после введения первой дозы и снижалась до поддер живаемого затем в опыте уровня 0,42+0,11 г/л в течение 40—60 мин. Первым поведенческим признаком интоксикации, наблюдавшимся на максимуме концентрации алкоголя, было выраженное и проходящее че рез 20—30 мин нарушение локомоции, которое могло не сопровождать ся нарушениями последовательности актов и эффективности пищедобы-вания. Время реализации цикла поведения по сравнению с КЭ в ЭЭ возрастало: с 7,87±1,98 до 11,24±3,80 с (р<0,001).
457
Как ошибки поведения рассматривались переходы от эффективной педали к неэффективной, остановки поведения — перерыв реализации цикла, проверки пустой кормушки без подхода к педали, подходы к пе дали без нажатия на нее. Число ошибок в ЭЭ достоверно превышало число ошибок в КЭ как после первого введения этанола, так и на под держивающих дозах. В КЭ и в ЭЭ кролики совершали разное число ошибок в правом и левом цикле, причем в ЭЭ выраженность этого раз личия увеличивалась (подробнее см. [4]).
Из 106 нейронов, активность которых была зарегистрирована в КЭ, 45% относились к группе вовлекающихся в обеспечение пищедобыва-тельного поведения, т. е. имеющих неизменно возникающие активации, приуроченные к определенному его этапу или этапам, а 55% —к груп пе нейронов, не вовлекающихся в обеспечение этого поведения, т. е. не имеющих таких активаций.
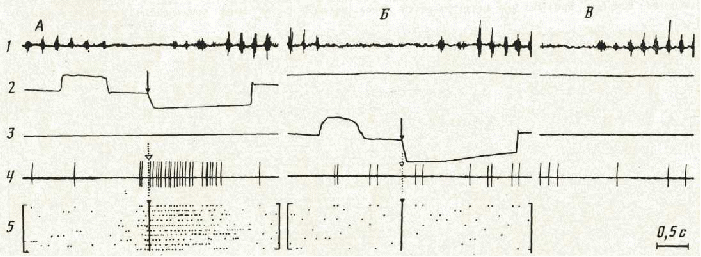
В соответствии с ранее описанными критериями выделения нейронов с разной поведенческой специализацией [2, 3] вовлекающиеся в обеспе чение поведения нейроны были разбиты на две подгруппы. Первая под группа— нейроны наиболее новых систем, сформированных при обуче нии животного инструментальному пищедобывательному поведению в экспериментальной камере. В КЭ первая подгруппа была представлена нейронами, активирующимися в актах достижения обеих кормушек, не смотря на то, что эти акты характеризовались оппонентными движения ми при реализации поведения у правой и левой стенок камеры. Другие нейроны этой подгруппы активировались в акте захвата пищи, но толь ко в определенных условиях: когда этот акт реализовался у правой или левой стенок камеры. На рис. 1 приведен пример активности такого нейрона, который активировался при наклоне, захвате и грызении пи щи в левой кормушке (А). Активация не появлялась в акте захвата пи щи из правой кормушки (Б), при наклонах головы вне кормушки, при захвате пищи, поданной экспериментатором с руки (В).
К первой подгруппе относились и нейроны, активирующиеся при подходах к обеим педалям (движение налево у правой стенки камеры и направо у левой) и/или нажатии на них (рис. 2, Б), а также активиру ющиеся в акте подхода и/или нажатия только на одну из педалей. Рис. 2, А иллюстрирует приуроченность активаций нейрона к этапам подхода и нажатия на левую педаль (1, 3). При подходе и нажатии на правую педаль активация не появлялась (2). Наконец, в КЭ у всех кро ликов были обнаружены «place-нейроны». Активации этих нейронов по являлись при пребывании кролика в определенном месте эксперимен тальной камеры. «Р1асе»-нейроны были отнесены к первой подгруппе потому, что их активации являются отражением «результативного» про странства, т. е. пространства, разбитого на участки в связи с поведен ческими актами, сформированными в конкретной среде [2].
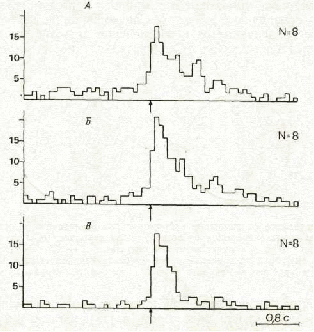
Вторая подгруппа — нейроны, специализированные относительно си стем, сформированных на этапах индивидуального развития, предшест вующих обучению животного инструментальному пищедобывательному поведению. В КЭ эта группа была представлена нейронами, которые активировались в связи с тем или иным движением тела и/или головы вне зависимости от того, в каком поведении это движение использова лось. На рис. 3 представлены гистограммы активности нейрона, активи рующегося в связи с движением головы налево в поведении у левой стенки (А, акт подхода к кормушке) и у правой стенки (Б, акт подхода к педали). Его активации возникали также в пассивно-оборонительном поведении, при смещениях головы животного налево экспериментато ром (рис. 3, В).
Нейроны первой подгруппы составляли 27% от числа нейронов, ак тивность которых была зарегистрирована в КЭ, а число нейронов вто рой подгруппы было меньшим—18%. При прохождении электрода че рез поперечник коры в ЭЭ удавалось выявлять в среднем 10,6 нейрона в треке — на треть меньше, чем в КЭ: 16,8 нейрона в треке (р<О,ОО1). Число не вовлекающихся в обеспечение поведения в ЭЭ нейронов не
Рис. 1. Пример активации нейрона, возникающей в акте захвата пищи из левой кормушки (ситуация ЭЭ). 1— ЭМГ т. masseter; 2 — актограмма поведения у левой стенки; 3 — у правой: отклонение кривых вверх — нажатие на педаль, вниз — наклон в кор мушку; 4 — импульсная активность нейрона; 5 — растры импульсной активности, построенные от момента пересечения носом плоскости отверстия кормушки (момент обозначен стрелками). А—В — пояснения в тексте
Рис. 2. Пример активаций, возникающих при подходе и нажатии только на левую педаль (А, ситуация КЭ) или при подходе и нажатии на обе педали (Б, ситуация ЭЭ). Все растры построены от момента начала нажатия на соответствующие педали (обозначен стрелками). Цифры слева от растров — последовательность реализации серий поведенческих циклов. На Л — активация возникает при подходе и нажатии на левую педаль — /, 3, но не на правую — 2; на Б— при подходе и нажатии на левую — 1 и правую — 2
педали
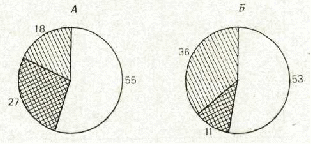
изменилось по сравнению с КЭ — 53% от общего числа проанализиро ванных в ситуации ЭЭ нейронов (n=110). Количественное же соотно шение нейронов, принадлежащих к первой и второй подгруппам груп пы вовлекающихся нейронов, резко изменилось: число нейронов первой группы уменьшилось с 27 до 11% (р <0,01), а число нейронов второй лодгруппы, увеличившись с 18 до 36% (р<0,01), стало достоверно (р < < 0,001) превышать число нейронов первой подгруппы (рис. 4).
Рис. 3. Пример активации, появляющейся при движении головы налево в разных по веденческих актах (ситуация ЭЭ). Гистограммы построены с использованием разрабо танного нами метода анализа видеозаписи [2] от момента начала движения налево (обозначен стрелками) при движении от педали к кормушке у левой стенки (А); от кормушки к педали у правой стенки (Б) и при смещениях головы животного налево экспериментатором (В). По оси ординат — число импульсов в канале (ширина кана ла— 80 мс), по оси абсцисс — время, с
Первая подгруппа в ЭЭ была представлена только нейронами, акти вирующимися в актах подхода и/или нажатия на педаль и в актах до стижения кормушки и/или захвата пищи из нее. «Place-нейроны» в ЭЭ обнаружены не были.
Количественное соотношение нейронов с «педальной» и «кормушеч-ной» специализацией в ЭЭ изменилось. В КЭ «кормушечные» нейроны составляли 8% от суммарного числа нейронов этих специализаций, а в ЭЭ — 42% (р<0,05, критерий Фишера).
Во второй подгруппе в ЭЭ кроме нейронов, составляющих ее в КЭ, обнаружены активирующиеся в акте захвата пищи нейроны, характер ные для антеролатеральной коры кролика [2, 3]. Эти нейроны, специа лизированные относительно систем, сформированных на ранних стади ях постнатального онтогенеза [2] и потому отнесенные ко второй под группе, в отличие от нейронов первой подгруппы, активирующихся в актах захвата пищи лишь в определенной поведенческой ситуации, да вали активации при достижении и захвате пищи в самых разных пове денческих ситуациях. Пример активности такого нейрона представлен на рис. 5. Он активируется не только при захвате пищи в обеих кор мушках (у правой — Б и левой — А стенок камеры), но и при захвате пищи, поданной экспериментатором с руки сверху (В).
461
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В экспериментах на кошках показано, что при острых введениях этанола в дозах, существенно превышающих использованные в наших экспериментах, не обнаруживается ни макро-, ни микроскопических по вреждений в корковых и подкорковых структурах [22]. Можно пола гать поэтому, что изменения нейронного обеспечения поведения, обна руживаемые при сравнении ЭЭ с КЭ, обусловлены обратимыми функ циональными изменениями активности.
Число нейронов, выявляемых в микроэлектродном треке, уменьша- ется в ЭЭ по сравнению с КЭ на 1/3. Соотношение же нейронов, вовлека ющихся и не вовлекающихся в обеспечение пищедобывательного пове дения, остается постоянным. Следовательно, имеет место уменьшение
Рис. 4. Количественное соотношение нейронов с разными типами специализации в КЭ и в ЭЭ. Цифры — число нейронов от общего числа нейронов (%), активность которых была зарегистрирована в соответствующей ситуации: А — КЭ, Б — ЭЭ. Незаштрихо-ванная часть — нейроны, не вовлекающиеся в обеспечение поведения. Штриховка пере секающимися линиями — нейроны первой подгруппы, параллельными — второй
на 1/3 каждой из групп. Изменение при этом соотношения нейронов пер вой и второй подгрупп говорит о том, что этанол при остром введении по-разному влияет на нейроны разных специализаций. Можно предпо ложить, что полученное изменение обусловлено либо увеличением чис ла активирующихся в поведении нейронов второй подгруппы и уменьше нием числа нейронов первой, либо уменьшением числа активирующихся нейронов первой подгруппы при неизменном числе нейронов второй подгруппы. Если умножить на 2/з число нейронов первой подгруппы (в процентах), обнаруженных в ЭЭ, то, естественно, получится число, еще более достоверно отличающееся (р <0,001) от числа этих нейронов в КЭ; при умножении на 2/з числа нейронов второй подгруппы достовер ного различия между ЭЭ и КЭ не обнаруживается. Таким образом, име ющиеся данные свидетельствуют в пользу второго предположения. Од нако они не позволяют отвергнуть первое. Во всяком случае, может быть принято следующее утверждение. При остром введении этанола имеет место уменьшение абсолютного числа активирующихся в пове дении нейронов первой подгруппы, т. е. нейронов наиболее новых сис тем. При анализе состава первой подгруппы в ЭЭ по сравнению с КЭ выявляется та же закономерность, что при сопоставлении первой и вто рой подгрупп. Процентное содержание нейронов, специализированных относительно систем, формируемых на начальных и завершающих ста диях обучения (см. методику), изменяется: первых — возрастает, вто рых — падает. Следовательно, нарушению пищедобывательного поведе ния при остром введении этанола соответствует уменьшение числа ак тивирующихся в поведении нейронов лимбической коры и изменение паттерна специализации нейронов [3] исследованной области, т. е. из менение количественного соотношения нейронов с разными типами спе циализации за счет исключения из обеспечения поведения части нейро нов наиболее новых систем. Сохранение выработанных поведенческих актов при резком увеличении числа ошибок может быть, по-видимому, объяснено тем, что использованные в настоящей работе дозы этанола
462
приводили к изменениям количественного соотношения нейронов разной специализации, но не к исчезновению активности всей совокупости ней ронов, специализированных относительно той или иной новой системы, сформированной в процессе обучения инструментальному поведению.
Данных для того, чтобы подтвердить или отвергнуть асимметрию паттерна специализации, соответствующую выявленной латерализации поведения, в настоящее время недостаточно.
Является ли избирательное угнетающее действие этанола на нейро ны новых систем закономерностью, общей для разных видов животных и форм поведения?
Рис. 5. Пример активации, возникающей при захвате пищи из левой (А) и правой кормушек (Б), а также пищи, поданной экспериментатором с руки (В) (ситуация ЭЭ). Растры построены от начала ЭМГ-активации т. masseter (обозначено стрелками), со ответствующей закрыванию рта при захвате пищи зубами. Треугольники — момент пе ресечения носом плоскости кормушки при наклоне (вершина вниз) и подъеме головы
(вершина вверх) из кормушки
Дж. Чапин и др. [11] показали, что введение этанола редуцирует имеющуюся в норме зависимость ответов нейронов первичной сомато-сенсорной коры крысы на стимуляцию их рецептивных полей от пове денческого контекста (покой, оборонительное поведение, ходьба). Ра нее [2] нами было экспериментально и теоретически обосновано сле дующее представление. Зависимость характеристик ответов нейронов на стимуляцию их рецептивных зон от того, в каком поведении эта стиму ляция имеет место, определяется тем, что в разных поведенческих ак тах эти нейроны согласуют свою активность с активностью разных со ставов нейронов. Разница составов связана, в частности, с различием наборов новых систем, реализуемых в разных поведенческих актах. Ес тественно ожидать, что угнетение нейронов новых систем должно при вести к уменьшению поведенческой модуляции активности нейронов, вовлекающихся в разное поведение, что и имело место в экспериментах Дж. Чапина и др., результаты которых, следовательно, являются аргу ментом в пользу утвердительного ответа на поставленный вопрос. В свя зи с этим уменьшение на 1/3 числа нейронов, не вовлекающихся в обес печение пищедобывательного поведения, можно предположительно объ яснить тем, что данная группа, по крайней мере частично, составлена из нейронов, специализированных относительно новых систем других форм поведения.
Особая чувствительность нейронов новых систем может рассматри ваться как механизм феноменов, выявляемых при исследовании влия ния острого введения этанола на память у людей и животных: этанол
463
действует на использование, приобретение и сохранение вновь выучи ваемого материала [10]. Следует отметить, однако, что действие остро го введения этанола на поведенческие акты определяется не только по следовательностью их формирования, но взаимодействием целого ряда факторов [4].
Многократно описанное в литературе явление диссоциации может выражаться как в невозможности реализовать в одном состоянии (на пример, на фоне действия алкоголя) поведение, сформированное в дру< гом состоянии (например, в норме), так и (для более простого поведе ния) в ухудшении характеристик его реализации [1]. А. А. Азарашвили [1] выдвинул следующую гипотезу, объясняющую феномены диссоции рованного обучения. При введении фармакологического вещества фор мируется новая «нейронная сеть», отличающаяся от той, которая обес печивала реализацию данного поведения в норме. Полученные нами факты подтверждают эту гипотезу. Для достижения результатов инст рументального пищедобывательного поведения при остром введении этанола формируется специальная интеграция, отличающаяся от ис ходной (КЭ) по числу и паттерну специализации нейронов. Различие сравниваемых интеграции, по-видимому, возрастает от более старых к более новым системам.
С позиций системных представлений о генезе импульсной активности она рассматривается как фактор, обеспечивающий достижение резуль тата функциональной системы, относительно которой нейрон специали зирован; при достижении результата активность нейрона прекращается [2, 9]. Следовательно, введение этанола приводит к эффекту, который для нейронов новых систем сопоставим с эффектом достижения резуль татов этих систем: прекращение активности, которое выражается в экс перименте с введением этанола в феномене уменьшения числа нейро нов. Подобная трактовка полученных данных не противоречит ни кон цепциям, рассматривающим этанол в качестве вещества-подкрепления (drug reinforcer) как для животных, так и для человека [19], ни пси хологическим представлениям о том, что на первой стадии алкоголиза ции самостоятельной потребности в алкоголе не существует; прием эта нола является способом удовлетворения других имеющихся у человека потребностей [6]. Кроме прямого действия на мембрану нейрона эта нол, изменяя активность других нейронов, а также практически все эта пы метаболизма [7], оказывает непрямое действие, которое определя ется особенностями медиаторных и рецепторных систем, кровоснабже ния, связей данной структуры и данного нейрона [16, 20, 24, 25]. До стижение результата, прекращающего активность нейрона, выступает для последнего как соответствующее изменение его «среды», зависящей от перечисленных выше (а возможно, и других) факторов. Можно пред положить, что этанол, оказывая прямое и непрямое действие на нейро ны всех специализаций, формирует такую «среду», которая оказывает ся по действию на активность нейронов новых систем в чем-то соответ ствующей «среде», формирующейся при достижении результатов этих систем в процессе реализации поведения.
Попытки выделения функциональных и структурных характеристик нейронов, определяющих то или иное действие этанола на активность данной клетки, привели к установлению целого ряда факторов: принад лежность к поли- или моносинаптической цепи, чувствительность к ме диаторам, свойства мембранных каналов и т. д. [11, 14, 17, 20, 25]. Ока зывается, однако, что критерий, например поли- или моносинаптическая связь нейрона с определенным входом, предсказывает эффективность действия этанола в одной структуре мозга, а в другой уже не работает [17]. Результаты настоящего исследования, демонстрирующие избира тельность действия этанола, зависящую от поведенческой специализа ции нейронов, позволяют предполагать, что то или иное действие этано ла на нейроны в конкретном поведении определяется специфическими наборами упомянутых выше и других характеристик, соответствующими специализации этих нейронов.
464
выводы
Нарушению инструментального пищедобывательного поведения у
кроликов, вызванному введением этанола, соответствует уменьшение
числа активных нейронов лимбической коры и изменение паттерна их
поведенческой специализации.Зависимость действия этанола от специализации нейрона прояв
ляется в том, что процентное содержание нейронов, обеспечивающих
реализацию наиболее новых систем, сформированных при обучении жи-
вотных инструментальному поведению, уменьшается, а нейронов, обес-
печивающих реализацию систем, сформированных на предыдущих эта-
пах индивидуального развития, увеличивается.Реализация инструментального пищедобывательного поведения в
норме и при введении этанола обеспечивается активностью разных на-
боров нейронов. Подобные различия, по-видимому, лежат в основе фе-
номенов диссоциации.
Список литературы
Азарашвили А. А. Диссоциированное обучение//Успехи физиол. наук. 1978. Т. 9.
№ 3. С. 95—114.Александров Ю. И. Психофизиологическое значение активности центральных и пе
риферических нейронов в поведении. М.: Наука, 1989. 207 с.Александров Ю. И., Гринченко Ю. В. Специализация нейронов моторной коры у
кроликов в норме и после разрушения зрительной коры//Журн. высш. нерв. деят.
1989. Т. 39. № 5. С. 914—923.Александров Ю. И., Гринченко Ю. В., Светлаев И. А., Абдрашитов О. X. К вопро-
су о факторах, определяющих влияние острого введения этанола на реализацию
поведения//Журн. высш. нерв. деят. 1989. Т. 39. № 6. С. 1149—1151.
5 Анохина И. П. Нарушения функции дофаминовой системы при алкоголизме//Био-логические основы алкоголизма. М.: МЗ СССР, 1984. С. 25—31.
Братусь Б. С. Психологический анализ изменений личности при алкоголизме. М.:
Изд-во МГУ, 1974. 95 с.Комиссарова И. А., Ротенберг Ю. С, Мастеропуло А. П. Механизмы действия эта
нола и подходы к коррекции обменных нарушений при хронической алкоголизации.
М., 1986. 74 с.Крыжановский Г. Н., Евсеев В. А. Нейропатофизиологический и нейроиммунопато-
логический подходы к пониманию механизмов и разработке принципов патогенети
ческой терапии алкоголизма//Вестн. АМН СССР. 1988. № 3. С. 10—14.Швырков В. Б. Нейрофизиологическое изучение системных механизмов поведения.
М.: Наука, 1978. 239 с.
Alkana R. L., Malcolm R. D. Comparison of the effects of acute alcohol intoxication
on behavior in humans and other animals//Animal models in alcohol research. L.:
Acad. Press, 1980. P. 193—268.Chapin J. K., Sorensen M. S., Woodward D. J. Acute ethanol effects on sensory res
ponses of single units in the somatosensory cortex of rats during different behavio
ral states//Pharmacol. Biochem. Behav. 1986. V. 25. No. 3. P. 607—614.Cloninger C. R. Neurogenetic adaptive mechanisms in alcoholism//Science. 1987.
V. 236. No. 4800. P. 410—416.Descarries L., Doucet G., Lemay B. et al. Structural basis of cortical monoamine fun-
ction//Neurotransmitters and cortical function. From molecules to mind. N. Y.; L.:
Plenum Press, 1988. P. 321—332.Faber D. S., Klee M. R. Actions of ethanol on neuronal membrane properties and
synaptic transmission//Alcohol and opiates. Neurochemical and behavioral mecha
nisms. N. Y.; L.: Acad. Press, 1977. P. 41—63.Grupp L. A., Perlanski E. Ethanol-induced changes in the spontaneous activity of
single units in the hippocampus of the awake rat: dose-response study//Neurophar-
macol. 1979. V. 18. No. 1. P. 63—70.Kalant H. Direct effects of ethanol on the nervous system//Federat. Proc. 1975.
V. 34. No. 10. P. 1930—1941.Kashii S., Ho J. Matsuoka I. et al. Effects of ethanol applied by electrosmosis on neu
rons in the lateral and medial vestibular nuclei//Japan. J. Pharmacol. 1984. V. 36.
No. 2. P. 153—159.Klemm W. R., Mallari С G., Dreyfus L. R. et al. Ethanol-induced regional and dose-
response differences in multiple-unit activity in rabbits//Psychopharmacol. 1976.
V. 49. No. 2. P. 235—244.Meisch R. A. Factors controlling drug reinforced behavior//Pharmacol. Biochem. Be
hav. 1987. V. 27. No. 2. P. 367—371.Mereu G., Gessa G. Low doses of ethanol inhibit the firing of neurons in the sub-
stantia nigra, pars reticulata: a GABAergic effect?//Brain. Res. 1985. V. 360. No. 1—
2. P. 325—330.Rogers J., Siggins G. R., Schutman J. A., Bloom F. E. Physiological correlates of
ethanol intoxication, tolerance, and dependence in rat cerebellar purkinje cells//Bra-
in Res. 1980. V. 196. No. 1. P. 183—198.
465
Sutko M. H., Weinberger N. М. Effects of ethanol on the cochlear nucleus and audi
tory cortex of the cat//J. Stud. Alcohol. 1979. V. 40. No. 9. P. 799—822.Vogt B. A., Sikes R. W., Swaldlow H. A., Weyand Th. G. Rabbit cinguiate cortex:
cytoarchitecture, physiological border with visual cortex, and different cortical con
nections of visual, motor, postsubicular and intracingulate origin//J. Compar. Neu-
rol. 1986. V. 248. No. 1. P. 74—94.Way tier M. J., Ono Т., Nolley D. Effects of ethyl alcohol on central neurons//Phar-
macol. Biochem. Behav. 1975. V. 2. No. 1. P. 499—506.Zornetzer S. F., Walker D. W., Hunter B. E., Abraham W. C. Neurophysiological
changes produced by alcohol//Biomedical processes and consequences of alcohol use.
Washington DC: U. S. Goverment Printing Office, 1982. P. 95—128.
Институт психологии Поступила в редакцию
АН СССР; Всесоюзный 27.1 V. 1989
научный центр Принята в печать
наркологии МЗ СССР, 15.XI.1989
Москва
EFFECT OF ACUTE ADMINISTRATION OF ETHANOL ON THE REALIZATION OF BEHAVIOUR AND ITS NEURONAL PROVISION
ALEXANDROV Yu. /., ORINCHENKO Yu. V., SVETLAEV I. A.
Institute of Psychology, USSR Academy of Sciences; All-Union Centre of Narcology, USSR Ministry of Health, Moscow
In experiments on rabbits trained to instrumental food procuring be haviour it was cleared up, which changes of activity of neurones of the limbic cortical area corresponded to disturbances of this behaviour (in crease in time of realization and in the number of errors) caused by intra-peritoneal injection of 12%' ethanol solution in a dose of 1 g/kg. In com parison with control (administration of isotonic solution), the number of active cells singled out in the microelectrode track was reduced by 1/3; the pattern of behavioural specialization of neurones involved in provision of the disturbed behaviour was changed. The content of neurones of the most recent systems formed during animals learning instrumental behaviour, decreased from 27 to 11%» and of neurones providing for realization of systems formed at previous stages of individual development increased from 18 to 36%.
466

Нравится материал? Поддержи автора!
Ещё документы из категории разное:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ