Клонирование животных
 Содержание
Содержание
Тезисы
Вступление
1. Из истории исследований по клонированию животных
2. Клонирование животных
3. Методы клонирования животных
3.1. Методы трансплантации ядер
3.2. SLIC (sequence and ligation-independent cloning) метод клонирования
3.3. Метод генетического перепрограммирования клеток кожи
4. Этические проблемы клонирования животных
5. Применения клонов животных
6. Эффективность клонирования животных
Выводы
Список использованной литературы
Тезисы
Клонирование, в биологии – это метод получения нескольких идентичных организмов путем бесполого (в том числе вегетативного) размножения.
Термин "клонирование" пришёл в русский язык из английского. Лишь немного изменив своё звучание и написание, он является аналогом английского clone, cloning. В самом же английском языке это слово стало употребляться (как биологический термин) менее 100 лет назад. Однако за этот небольшой для жизни слова срок оно уже успело несколько раз поменять своё значение.
Создавать животных и растения с заданными качествами всегда было чем-то чрезвычайно заманчивым так как это означало создать организмы уникальнейшие и нужнейшие, устойчивые к болезням, климатическим условиям, дающие достаточный приплод, необходимое количество мяса, молока, плодов, овощей и прочих продуктов. Использование таких технологий клонирования предполагает уникальную возможность получать фенотипически и генетически идентичные организмы, которые могут быть использованы для решения различных теоретических и прикладных задач, стоящих перед биомедициной и сельским хозяйством. В частности, использование клонирования могло бы способствовать изучению проблемы тотипотентности дифференциированных клеток, развития и старения организмов, злокачественного перерождения клеток. Благодаря технологиям клонирования предполагается появление ускоренной генетической селекции и тиражирования животных с исключительными производственными показателями. В сочетании с трансгенозом клонирование животных открывает дополнительные возможности для производства ценных биологически активных белков для лечения различных заболеваний животных и человека. Клонирование животных возможно позволит проводить испытания медицинских препаратов на идентичных организмах.
Все клетки организма животных несут одинаковую генетическую информацию. Однако в процессе морфогенеза соматические клетки дифференцируются, в результате чего часть генома репрессируется. Чем выше уровень специализации клеток, тем меньше их тотипотентность. Эта закономерность была установлена в экспериментах по пересадке ядер.
Вступление
"Клонирование" получение потомков, являющихся точной генетической копией организма. Совокупность таких потомков-копий, происходящих от одного организма, называют клоном. Организмы в пределах каждого клона характеризуются одинаковой фенотипической однородностью и идентичным генотипом.
Термин "клон" был впервые использован в 1903 году Веббером (Webber, Германия) применительно к растениям, размножаемым вегетативно, и означал, что дочерние растения клона генетически идентичны материнскому. В настоящее время разработки в области генной инженерии позволяют клонировать не только микроорганизмы и растения, но и животных. Впервые трансплантацию ядер соматических клеток зародышей в энуклеированные клетки лягушки осуществили американские исследователи Р. Бриггс и Т. Кинг в 1952 году. Ученые, пользуясь микропипеткой, удаляли ядра из яйцеклеток шпорцевой лягушки, а вместо них пересаживали ядра клеток эмбрионов, находящихся на разных стадиях развития. Проведенные исследования показали, что ядра ранних эмбрионов в стадии поздней бластулы и даже ранней гаструлы обладают тотипотентностью и обеспечивают нормальное развитие эмбрионов. Если брать ядра из клеток зародыша на ранней стадии его развития - бластуле, то примерно в 80% случаев зародыш благополучно развивается дальше и превращается в нормального головастика. Если же развитие зародыша, донора ядра, продвинулось на следующую стадию - гаструлу, то лишь менее чем в 20% случаев оперированные яйцеклетки развивались нормально. При пересадке ядер из более дифференцированных клеток (мезодермы и средней кишки) поздней гаструлы у эмбрионов наблюдалось недоразвитие и даже отсутствие нервной системы. После пересадки ядра из клеток более позднего развития яйцеклетки вообще не развивались.
Из истории исследований по клонированию животных
Возможность клонирования животных доказал Дж. Гердон, английский биолог, который первым сумел получить клонированные эмбрионы шпорцевых лягушек. Он выжигал ультрафиолетом ядра икринок и затем подсаживал в них ядра, выделенные из клеток эпителия головастиков этого вида. Большая часть полученных таким образом икринок погибала, и лишь совсем маленькая их доля (2,5%) развивалась в головастиков. Взрослых лягушек получить таким образом не удавалось. Тем не менее это был успех, и результаты опытов Гердона попали во многие учебники и руководства по биологии. В 1976 г. Гердон и его соавтор Р. Ласки публикуют работу, в которой описывают опыты с ядрами, выделенными из клеток почек, кожи и легкого уже взрослых шпорцевых лягушек. Исследователи сначала подращивают эти клетки вне организма (in vitro), а затем вводят их ядра в безъядерные икринки. Четверть таких икринок начинает делиться, но вскоре замирает на одой из стадий развития. Тогда ученые выделяют ядра полученных эмбрионов и снова подсаживают их в лишенные собственных ядер икринки... В результате целой серии подобных пересадок на свет наконец-то появляется несколько головастиков. Хотя эксперименты Гердона и его последователей показали принципиальную возможность получения серийных клонов амфибий, появляющиеся на свет головастики упорно не желали превращаться во взрослых лягушек. Вопрос, таким образом, по-прежнему заключался в том, можно ли вырастить из одной специализированной клетки его тела взрослое позвоночное животное. Опыты на амфибиях давали отрицательный результат, но ученые не прекращали исследований в этой области.
Более широкие исследования, охватывающие не только амфибий, но и рыб, а также дрозофил, в 1962 г. были начаты английским биологом Дж. Гордоном. Он первым в опытах с южноафриканскими жабами Xenopus laevis) в качестве донора ядер использовал не зародышевые клетки, а уже вполне специализировавшиеся клетки эпителия кишечника плавающего головастика.
Затем Гердон вместе с Ласки (1970) стали культивировать in vitro (вне организма в питательной среде) клетки почки, легкого и кожи взрослых животных и использовать уже эти клетки в качестве доноров ядер. Примерно 25% первично реконструированных яйцеклеток развивались до стадии бластулы. При серийных пересадках они развивались до стадии плавающего головастика. Таким образом было показано, что клетки трех разных тканей взрослого позвоночного (X. laevis) содержат ядра, которые могут обеспечить развитие по крайней мере до стадии головастика.
В свою очередь Ди Берардино и Хофнер (1983) использовали для трансплантации ядра неделящихся и полностью дифференцированных клеток крови - эритроцитов лягушки Rana pipiens. После серийной пересадки таких ядер 10% реконструированных яйцеклеток достигали стадии плавающего головастика. Эти эксперименты показали, что некоторые ядра соматических клеток способны сохранять тотипотентность.
Причины, по которым ядра клеток взрослых животных и даже поздних эмбрионов остаются тотипотентными, пока точно не установлены. Решающую роль играет взаимодействие ядра и цитоплазмы. Содержащиеся в цитоплазме животных вещества принимают участие в регулировании экспрессии клеточного генов ядра[5].
Работы М. ди Бернардино и Н. Хоффера показали, что цитоплазма ооцитов амфибий содержит факторы, восстанавливающие тотипотентность ядер дифференцированных соматических клеток. Эти факторы реактивируют репрессированные участки генома.
В 1985 г. была описана технология клонирования костных рыб, разработанная советскими учеными Л.А. Слепцовой, Н.В. Дабагян и К.Г.Газарян. Зародыши на стадии бластулы отделяли от желтка. Ядра клеток зародышей впрыскивали в цитоплазму неоплодотворенных икринок, которые начинали дробиться и развивались в личинки. Эти эксперименты показали, что потеря ядром тотипотентности в процессе онтогенеза связана не с утерей генов, а их репрессией. При культивировании соматических клеток in vitro частота тотипотентности ядер увеличивается. Генетический механизм стабильной репрессии генома дифференцированных клеток не выяснен, способы восстановления тотипотентности не разработаны, поэтому в основном ведется клонирование путем трансплантации ядер эмбриональных клеток.
Пересадки ядер у млекопитающих начались позднее, в 80-х годах. Это было связано с техническими трудностями, так как зигота млекопитающих имеет небольшие размеры. Например, диаметр зиготы мыши приблизительно 60 мкм, а диаметр оплодотворенной яйцеклетки лягушки около 1200 мкм, т.е. в 20 раз больше[26].
Несмотря на перечисленные трудности, первые сообщения о получении клонов мышей, идентичных донору, появились уже в 1981 году. В качестве донора были использованы эмбриональные клетки одной из линий мышей, взятые на стадии бластоцисты. Достоверность полученных данных вначале была поставлены под сомнение, так как воспроизвести результаты проведенных экспериментов в других лабораториях не удавалось, однако пару лет спустя Дж. Мак Грат и Д. Солтер также достигли успеха. В этих экспериментах клоны мышей удавалось получить лишь в том случае, если трансплантировали ядра эмбрионов на стадии не позднее 2 бластомеров. Было показано, что ядра 8-клеточных зародышей и клеток внутренней клеточной массы бластоцисты не обеспечивают развитие in vitro реконструированных яйцеклеток даже до стадии морулы, которая предшествует стадии бластоцисты. Небольшая часть (5%) ядер 4-клеточных зародышей дает возможность развиваться только до стадии морулы. Эти и многие другие данные показывают, что в эмбриогенезе у мышей клеточные ядра рано теряют тотипотентность, что связано очевидно, с очень ранней активацией генома зародыша - уже на стадии 2-х клеток. У других млекопитающих, в частности, у кроликов, овец и крупного рогатого скота, активация первой группы генов в эмбриогенезе происходит позднее, на 8-16-клеточной стадии. Возможно поэтому первые значительные успехи в клонировании эмбрионов были достигнуты на других видах млекопитающих, а не на мышах. Тем не менее, работы с мышами, несмотря на их непростую судьбу, значительно расширили наши представления о методологии клонирования млекопитающих.
В начале пути
1883 — Открытие яйцеклетки немецким цитологом Оскаром Гертвигом.
1943 — Журнал Science сообщил об успешном оплодотворении яйцеклетки "в пробирке".
1977 — Профессор зоологии Оксфордского университета Дж. Гордон клонирует более полусотни лягушек.
1978 — Рождение в Англии Луизы Браун, первого ребёнка "из пробирки".
1985 — 4 января в одной из клиник северного Лондона родилась девочка у миссис Коттон — первой в мире суррогатной матери (зачата не из яйцеклетки миссис Коттон).
1987 — Специалисты Университета имени Дж. Вашингтона, использовавшие специальный фермент, сумели разделить клетки человеческого зародыша и клонировать их до стадии тридцати двух клеток (бластов, бластомеров).
Дж. Гордон
Первые успешные опыты по клонированию животных были проведены в середине 1970-х годов английским эмбриологом Дж. Гордоном (J. Gordon) в экспериментах на амфибиях, когда замена ядра яйцеклетки на ядро из соматической клетки взрослой лягушки привела к появлению головастика. Это показало, что техника трансплантации ядер из соматических клеток взрослых организмов в энуклеированные ооциты позволяет получать генетические копии организма, послужившего донором ядер дифференциированных клеток. Результат эксперимента стал основанием для вывода об обратимости эмбриональной дифференцировки генома по крайней мере у земноводных[18].
2. Клонирование животных
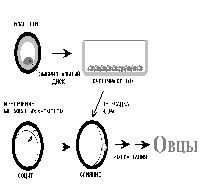
В своем эксперименте Кэмпбелл и его коллеги извлекли из эмбриона овцы на ранней стадии развития (на стадии эмбрионального диска) клетку и вырастили культуру клеток, то есть добились того, что клетка размножилась в искусственной питательной среде. Полученные генетически идентичные клетки (клеточная линия) сохранили тотипонентность. Затем ученые взяли яйцеклетку овцы-реципиента, тщательно удалили из нее весь хромосомный материал и добились ее слияния с тотипотентной клеткой из культуры. Полученные синтетические эмбрионы выращивали до стадии морулы-бластулы, а затем имплантировали в матку овцы. В результате удалось вырастить нескольких нормальных ягнят, которые были генетически идентичны.
Рис. 1. Методика, с помощью которой Кэмпбелл и его коллеги клонировали овец.
Из клеток эмбрионального диска получили устойчивые культуры клеток. Из ооцитов-реципиентов удаляли часть цитоплазмы вместе с метафазной пластинкой и индуцировали слияние таких безъядерных
ооцитов с клеткой из тотипотентной клеточной линии. Полученные таким образом эмбрионы временно помещались в овцу-реципиента. через неделю проверяли уровень их развития. Наконец, морулы и бластоцисты имплантировались другим овцам, где и проходил весь онтогенез.
В принципе, после того, как получена устойчивая линия тотипонентных клеток, ничто не мешает вносить в них генетические изменения. Например, перестраивая или удаляя отдельные гены, можно создавать трансгенные линии овец и других сельскохозяйственных животных. Однако прежде чем эта технология найдет практическое применение, предстоит решить еще множество проблем.
Пока число клонированных животных очень мало по сравнению с числом исходных эмбрионов, из клеток которых удавалось получить культуру. Многие клетки погибали, не успев достичь стадии бластоцисты. Не ясно, вызван ли высокий процент неудач разнообразными вредными факторами, воздействующими на клетку при манипуляциях с нею, или гетерогенностью самой клеточной линии. Последнее менее вероятно, поскольку процент успешных случаев не меняется при пересевах культуры. Для прояснения этого вопроса необходимо исследовать другие тотипотентные клеточные линии.
Результативность пересадки ядра в яйцеклетку и ее последующее благополучное развитие зависит от адекватного перепрограммирования ядра донора. Макромолекулы (белки и транспортная РНК) ооцита отвечают за его развитие только в течение сравнительно короткого времени (между двумя клеточными делениями), и чем этот период короче, тем меньше остается времени для перепрограммирования. Клетки более зрелых эмбрионов требуют большего времени для перепрограммирования, поэтому вероятность успеха при их использовании снижается. Определенную роль играет также совместимость ядра донора и цитоплазмы реципиента, все еще слабо изученная.
Успех пересадки клеточных ядер связан по крайней мере с двумя факторами. Во-первых, овулировавшие ооциты являются лучшими реципиентами, чем зиготы, либо потому, что у неоплодотворенных яйцеклеток остается больше времени для перепрограммирования, либо потому, что их цитоплазма является более подходящей. Возможно, в цитоплазме ооцита есть элементы, необходимые для перестройки хромосом и активации генома и исчезающие после оплодотворения либо потому, что они каким-то образом связаны с реплицирующейся ДНК, либо в результате запрограммированного распада. Во-вторых, клетки с ядрами донора, взятыми на стадиях G1 или G0 клеточного цикла, развиваются гораздо лучше, чем клетки с ядрами со стадий S или G2. Интуитивно это кажется понятным, ведь перепрограммировать открытый реплицирующийся геном проще.
Клонирование животных возможно с помощью экспериментальных манипуляций с яйцеклетками (ооцитами) и ядрами соматических клеток животных in vitro и in vivo подобно тому, как в природе появляются однояйцевые близнецы. Клонирование животных достигается в результате переноса ядра из дифференцированной клетки в неоплодотворённую яйцеклетку, у которой удалено собственное ядро (энуклеированная яйцеклетка) с последующей пересадкой реконструированной яйцеклетки в яйцевод приёмной матери. Однако долгое время все попытки применить описанный выше метод для клонирования млекопитающих были безуспешными. Значительный вклад в решение этой проблемы был сделан шотландской группой исследователей из Рослинского института и компании "PPL Therapeuticus" (Шотландия) под руководством Яна Вильмута (Wilmut). В 1996 году появились их публикации по успешному рождению ягнят в результате трансплантации ядер, полученных из фибробластов плода овцы, в энуклеированные ооциты. [2] В окончательном виде проблема клонирования животных была решена группой Вильмута в 1997, когда родилась овца по кличке Долли — первое млекопитающее, полученное из ядра взрослой соматической клетки: собственное ядро ооцита было заменено на ядро клетки из культуры эпителиальных клеток молочной железы взрослой лактирующей овцы. [3] В дальнейшем были проведены успешные эксперименты по клонированию различных млекопитающих с использованием ядер, взятых из взрослых соматических клеток животных (мышь, коза, свинья, корова), а также взятых у мёртвых, замороженных[4] на несколько лет, животных. Появление технологии клонирования животных вызвало не только большой научный интерес, но и привлекло внимание крупного бизнеса во многих странах. Подобные работы ведутся и в России, но целенаправленной программы исследований не существует. В целом технология клонирования животных ещё находится в стадия развития. У большого числа полученных таким образом организмов наблюдаются различные патологии, приводящие к внутриутробной гибели или гибели сразу после рождения.
В апреле 2008 года Южнокорейские таможенники приступили к дрессировке семи щенков, клонированных из соматических клеткок лучшего корейского розыскного пса породы канадский лабрадор-ретривер. По мнению южнокорейских ученых, 90 % клонированных щенков будут удовлетворять требованиям для работы на таможне, тогда как лишь менее 30 % обычных щенков проходят тесты на профпригодность.
Клонирование с целью воссоздания вымерших видов
Клонирование может быть использовано для воссоздания естественых популяций животных, вымерших по вине человека. Несмотря на наличие определённых проблем и трудностей, первые результаты в данном направлении уже имеются.
Клонирование бантенгов
В 2004 году на свет появилась пара бантенгов (диких быков, обитавших в Юго-Восточной Азии), клонированных из клеток животных, умерших более 20 лет назад. Два бантенга были клонированы из уникального "замороженного зоопарка" Сан-Диего, созданного еще до того, как люди поняли, что клонирование вообще возможно. Произведшая клонирование американская компания Advanced Cell Technology сообщила, что в нем использовались клетки животных, которые умерли в 1980 году, не оставив потомства.
Бантенгов клонировали, перенеся их генетический материал в пустые яйцеклетки обычных домашних коров; из 16 зародышей до рождения дожили только два. [7] [8]
Императорский дятел
В последний раз императорского дятла видели в Мексике в 1958 году. С тех пор орнитологи пытаются найти следы этой популяции, но безуспешно. Около десяти лет назад появились даже слухи, что птица еще живет на планете, но и они не подтвердились.
Зато в музеях остались чучела птицы. Научный сотрудник Дарвиновского музея Игорь Фадеев считает, что если операцию по выделению ДНК провести со всеми чучелами, которые находятся в разных странах мира, то дятла можно будет воскресить. В разных музеях мира на сегодняшний день осталось лишь десять чучел императорского дятла.
Если проект увенчается успехом, то в недалеком будущем на нашей планете, возможно, вновь появится императорский дятел. В Государственном Дарвиновском музее уверены, что последние методы молекулярной биологии позволяют выделить и воспроизвести ДНК этих птиц. [9]
Дронт
В июне 2006 года голландские учёные обнаружили на острове Маврикий хорошо сохранившиеся останки дронта - вымершей исторически недавно (в XVII веке) нелетающей птицы. Ранее наука не располагала останками птицы, в исчезновении которой, как всегда, виноват человек. Но теперь появилась определенная надежда на "воскресение" удивительного представителя пернатых. [10]
Клонирование гигантских птиц
Планы по клонированию исчезнувших гигантских птиц были поставлены под сомнение в результате исследований учёных Оксфордского университета. Выделив участки ДНК из останков вымерших птиц, ученые обнаружили, что их генетический материал настолько разрушен, что современная технология не позволяет провести полноценное клонирование. Цель научных работ состояла в возрождении вымерших несколько веков назад новозеландского страуса Моа, а также Мадагаскарского эпиорниса (птицы-слона).
Образцы ДНК были взяты из фрагментов тканей, сохранившихся в музеях. Однако ученые не смогли получить достаточную по своей длине цепочку ДНК, чтобы провести клонирование. Тем не менее, некоторые ученые считают, что в ближайшие годы будет разработана технология восстановления утраченных частей ДНК, путем вшивания туда "заплат" из ДНК близкородственных видов.
1970 - успешное клонирование лягушки[12]
1985 - клонирование костных рыб[13]
1996 — овечка Долли.
1997 — первая мышь.[14].
1998 — первая корова[15].
1999 — первый козёл[16].
2001 — первая кошка[17].
2002 — первый кролик[18].
2003 — первые бык[19], мул[20], олень[21].
2004 — первый опыт клонирования с коммерческими целями (кошки).[22]
2005 — первая собака (афганская борзая по кличке Снуппи).[23]
2006 — первый хорёк
2007 - вторая собака [24]
2008 — третья собака (лабрадор по кличке Чейс). Клонирована по государственному заказу[25]. Начало коммерческого клонирования собак[26]
Методы клонирования животных
Последние десятилетия XX века ознаменовались бурным развитием одной из главных ветвей биологической науки -- молекулярной генетики. Уже в начале 70-х годов ученые в лабораторных условиях начали получать и клонировать рекомбинантные молекулы ДНК, культивировать в пробирках клетки и ткани растений и животных. Возникло новое направление генетики генетическая инженерия. На основе ее методологии начали разрабатываться различного рода биотехнологии, создаваться генетически измененные организмы (ГМО). Появилась возможность генной терапии некоторых заболеваний человека, а последнее десятилетие XX века ознаменовалось еще одним важным событием -- достигнут огромный прогресс в клонировании животных из соматических клеток.
Особенно большой резонанс у мировой общественности получили исследования шотландских ученых из Рослинского Университета, которым удалось из клетки молочной железы беременной овцы получить генетически точную ее копию. Клонированная овца по кличке Долли нормально развивалась и произвела на свет сначала одного, а затем еще трех нормальных ягнят. Вслед за этим появился ряд новых сообщений о воспроизведении генетических близнецов коров, мышей, коз, свиней из соматических клеток этих животных. У приматов, в частности, у обезьян пока не удалось получить клоны с использованием клеток взрослого организма, плода или даже эмбриональных стволовых клеток.
Тем не менее работы в этом направлении активно ведутся. В прошлом году появилось сообщение о клональном размножении потомства приматов путем деления зародыша. Американским исследователям удалось получить генетически идентичные эмбрионы обезьяны резус путем разделения бластомеров зародыша на стадии деления. Из эмбриона родилась вполне нормальная обезьянка Тетра.
Такой тип клонирования обеспечивает генетически идентичное потомство, и в результате можно получить двойню, тройню и более генетических близнецов. Это позволяет проводить теоретические исследования по эффективности новых методов терапии тех или иных заболеваний, появляется возможность повторять научные эксперименты на абсолютно генетически идентичном материале. Имплантируя зародыши последовательно одной и той же суррогатной самке, можно исследовать влияние ее организма на развитие плода [31].
Разработанные методы клонирования животных пока еще далеко не совершенны. В процессе экспериментов наблюдается высокая смертность плодов и новорожденных. Еще не ясны многие теоретические вопросы клонирования животных из отдельной соматической клетки.
Тем не менее успех, достигнутый в клонировании овцы и обезьян, показал теоретическую возможность создания генетических копий также человека из отдельной клетки, взятой из какого-либо его органа. Многие ученые с энтузиазмом восприняли идею клонирования человека.
3.1 Методы трансплантации ядер
В нашей стране Б.В. Конюховым и Е.С. Платоновым в 1985 г. был разработан метод менее травматического переноса ядер методом микроманипуляции. Он протекает в два этапа: сначала тонкой микропипеткой прокалывают зоны пеллюцида и плазматической мембраны и извлекают пронуклеусы, а затем другой пипеткой, большего диаметра (12 мкм) в то же отверстие вводят диплоидное ядро донора. В этом случае меньше травмируется цитоплазма зиготы и транспортируемое ядро донора.
Трансплантация ядер может осуществляться и другим способом, с использованием цитохалазинов (веществ, синтезируемых грибами).
Цитохалазин В разрушает структуру микрофиламентов и способствует уникальному расположению ядра. Ядро остается соединенным с клеткой тоненьким стебельком цитоплазмы. При центрифугировании этот мостик разрывается, образуются безъядерные клетки (цитопласты) и кариопласты, представляющие собой ядра, окруженные тонким слоем цитоплазмы и цитоплазматической мембраной. Цитопласты отделяют от интактных клеток в градиенте плотности. Они сохраняют способность прикрепляться к поверхности культурального сосуда и могут быть использованы для слияния с кариопластами других клеток с целью получения жизнеспособной клетки [14][12].
Методы выделения кариопластов несколько сложнее и включают в себя ряд операции по центрифугированию, разделению в градиенте плотности и т.д. В некоторых случаях к смеси клеток и кариопластов добавляют частицы тантала диаметром 1 – 3 мкм. Они проникают в клетки и никогда в кариопласт, поэтому более тяжелые клетки осаждаются быстрее кариопластов.
Цитопласты содержат все виды органелл, присущие нормальной клетке, сохраняют способность прикрепляться к субстрату, образовывать складчатую мембрану, передвигаться, осуществлять пиноцитоз.
Кариопласты окружены тонким слоем цитоплазмы (около 10% от всей клеточной цитоплазмы), содержат компактный эндоплазматический ретикулум, несколько митохондрий и рибосом. У некоторых клеточных линий 1/10 кариопластов способна восстановить весь утраченный объем цитоплазмы и восстановиться в жизнеспособные клетки.
Для реконструкции клеток суспензию кариопластов в солевом буфере добавляют к монослою культуры цитопластов из пропорции 100 кариопластов на 1 цитопласт. Цитопласты должны быть уже покрыты инактивированными вирусными частицами. Инкубируют при температуре 4оС 45 минут, а затем еще 45 минут при температуре 37оС. Отмывают раствором Эрла для удаления не слившихся кариопластов [8].
3.2 SLIC (sequence and ligation-independent cloning) метод клонирования
Стив Элледж и неутомимая Мами Ли в очередной раз порадовали научное сообщество оригинальным методом клонирования. Новый метод получил название SLIC (sequence and ligation-independent cloning). Новинка является модификацией известного метода LIC (ligation-independent cloning) - клонирования без использования лигазы. Для того чтобы вставить фрагмент ДНК в вектор при помощи классического метода LIC, достаточно смешать вектор и вставку, на концах которых расположены протяженные одноцепочечные участки, комплементарные друг другу. При этом вставка "прилипает" к вектору, образуя рекомбинантную плазмиду с никами в обеих цепях. Полученной плазмидой трансформируют Е.coli, система репарации которой восстанавливает нормальную структуру плазмиды.
Метод SLIC - это то же самое, что и LIC с единственной разницей: "слипание" вектора и вставки проводят в присутствии белка RecA. Эта незначительная модификация метода позволяет добиться достаточно высокого выхода (1 нг. вектора способен дать 3900 трансформантов) а также упростить саму процедуру клонирования. Так если для классического метода LIC необходимо точно подогнать размер одноцепочечных участков у вектора и вставки (чтобы в итоге на стыке вектора и вставки получились ники), то метод SLIC допускает наличие протяженных гэпов [19].
Фактор RecA- один из ключевых факторов репарации и рекомбинации E.coli. Связываясь с одноцепочечным участком ДНК, RecA стимулирует процесс "strand exchange" (в ходе этого процесса одноцепочечный участок одной молекулы ДНК встраивается в гомологичный двухчепоченый участок другой молекулы ДНК, образуя D-петлю). Очевидно, добавление RecA in vitro на шаге клонирования позволяет E.coli эффективнее репарировать плазмиду in vivo.
Для успешного клонирования необходимо наличие 30-ти нуклеотидных участков гомологии по краям вектора и вставки. Получать одноцепочечные участки предлагается с помощью T4 ДНК-полимеразы без добавления нуклеотидов. При помощи SLIC в один вектор можно запихнуть сразу 5 вставок в одну стадию без снижения выхода. Наконец, высокий выход позволяет использовать метод SLIC при клонировании библиотек [7].
3.3 Метод генетического перепрограмирования клеток кожи
Разработан новый метод клонирования – менее трудоемкий, чем способ, благодаря которому была создана овечка Долли. В связи с этим возникли опасения, что однажды он будет использован для обработки эмбрионов человека, дабы формировать детей "по заказу".
Ученые, благодаря этому методу получившие мышат из клеток кожи взрослых особей, обнаружили: такая технология намного более эффективна, чем способ создания Долли, а побочных эффектов у нее меньше – следовательно, она лучше подходит для использования применительно к человеку.
Для клонирования мышей ученые вводили клетки кожи, взятые у взрослой особи, в ткани эмбриона на ранней стадии развития, полученного путем экстракорпорального оплодотворения (ЭКО). Некоторые из детенышей оказались частичными клонами особей-доноров, а некоторые, как и Долли, стопроцентными.
Однако, в отличие от "метода имени Долли", этот способ настолько прост и эффективен, что возникли опасения: в клиниках, где практикуется ЭКО человека, им могут воспользоваться для помощи бесплодным супружеским парам, которые мечтают о полностью "своем" в биологическом отношении ребенке[11].
Метод предполагает генетическое перепрограммирование клеток кожи, в результате которого они возвращаются в квазиэмбриональное состояние. В прошлом году, когда эта революционная методика впервые была применена к клеткам кожи человека, Католическая церковь и президент Джордж Буш высоко оценили ее как нравственно-приемлемый способ получения эмбриональных стволовых клеток, не сопряженный с необходимостью создавать или уничтожать человеческие эмбрионы [13].
Однако тот же метод уже используется в иных целях – для воспроизводства потомства лабораторных мышей, которое является либо стопроцентными клонами, либо генетическими "химерами" взрослых мышей, клетки кожи которых подверглись перепрограммированию.
Эксперименты на мышах показали, что в принципе теперь возможно взять клетку кожи человека, перепрограммировать ее для возврата в эмбриональное состояние, а затем ввести ее в эмбрион человека на ранней стадии. В результате получится ребенок, обладающий некоторыми общими генами не только с родителями эмбриона, но и с человеком, у которого были взяты клетки кожи.
Такой ребенок является химерой – генетической "помесью" двух или большего числа особей – так как некоторые из его клеток происходят от эмбриона, а другие – от клетки кожи. Фактически у такого ребенка будет три биологических родителя. Известны химеры человека, возникающие в естественных условиях – когда в матке соединяются два эмбриона. Часто подобные люди являются совершенно нормальными и здоровыми. По словам доктора Ланцы, нет причин предполагать, что люди-химеры, созданные при помощи нового метода, будут нездоровы.
Более того, эксперименты на мышах показали, что возможно создавать полные клоны – детенышей, которые на 100% идентичны взрослой особи в генетическом плане. Этого удалось достигнуть, используя разновидность дефективных эмбрионов мышей с четырьмя наборами хромосом вместо нормального числа – двух.
Этот "тетраплоидный" эмбрион, развиваясь, превращался исключительно в плаценту плода; когда же в него ввели перепрограммированную клетку кожи, остальная часть плода развилась из этой единственной клетки и сделалась стопроцентным клоном взрослой особи, кожа которой использовалась [15].
Никто из ученых, разрабатывающих методы перепрограммирования клеток для производства индуцированных плюрипотентных стволовых клеток (induced pluripotent stem, сокращенно iPS) – так называют эмбриональные клетки – не планирует применять их в репродуктивной медицине человека. Главная цель ученых – наладить производство стволовых клеток для терапевтического лечения таких заболеваний, как болезнь Паркинсона, болезнь Альцгеймера и инсульт [3].
Этические проблемы клонирования животних
Члены Европарламента на пленарном заседании ассамблеи в Страсбурге проголосовали "широким большинством голосов" против клонирования животных в продовольственных целях, сообщил РИА Новости представитель пресс-службы Европарламента.
На сегодняшний день нигде в мире продукты из мяса животных-клонов не продаются, однако, по мнению экспертов, они могут появиться на продовольственных рынках к 2010 году, в частности, в США. В марте комитет по безопасности продуктов питания при правительстве Японии сделал заключение о том, что мясо клонированных коров и свиней безопасно.
Европейское агентство по продовольственной безопасности (EFSA) в январе прошлого года обнародовало предварительное заключение, согласно которому, мясо и молоко клонированных животных пригодно в пищу. Эксперты EFSA, проводившие исследование по поручению Еврокомиссии, считают "очень маловероятным, что существует какое-либо отличие в плане продовольственной безопасности" между мясом и молоком животных-клонов и их сородичей, выращенных традиционным путем.
Независимая Европейская экспертная группа по этике в области науки и новых технологий (EGE), со своей стороны, высказала сомнения в оправданности возможности потребления человеком мяса и молока животных-клонов. Эксперты EGE исходят из того, что подсадка эмбрионов клонов самкам домашних животных является болезненным процессом и зачастую чревата их гибелью, а сами клонированные животные преждевременно стареют и очень уязвимы в плане здоровья.
Решение о разрешении к продаже мяса и молока животных-клонов надлежит принимать Еврокомиссии совместно со странами-членами ЕС [24].
5. Применения клонов животных
Клоны не всегда выглядят одинаково. Хотя у клонов один и тот же генетический материал, окружающая среда так же играет огромную роль в том, как приспособится к ней организм клонированного существа. К примеру, первая кошка, которая была клонирована, ее звали Сиси, была каленкорской породы и была совсем не похожа на свою мать (донора) [21].
Репродуктивное клонирование может позволять исследователям клонировать животных с потенциальной выгодой для областей медицины и сельского хозяйства. Например, те же самые Шотландские исследователи, которые клонировали Долли, клонировали другую овцу, которая была генетически модифицирована, чтобы давать молоко, которое содержит человеческую основу белка для крови . Мы надеемся, что в дальнейшем этот белок может отбираться из молока и подаваться человеку в чистом виде, это очень поможет людям, у которых низкая свертываемость крови. Так же можно использовать животных для того чтобы тестировать на них новые виды лекарств и обычную продукцию, предназначенную для человека. Большое преимущество использования клонированных животных для проверки на таблетки состоит в том, что все они являются генетически идентичными, что означает, что их реакция на таблетки должна быть боле менее сходной, чем у животных с различным генетическим набором.
Другой причиной для клонирования может служить то, что существуют популяции животных, которые стоят на грани вымирания. В 2001 году именно по этой причине ученые произвели первого клона, подвергнутого опасности вымирания - азиатского вола.
Детеныш, который развивался в матке у своей мамы-заместителя погиб всего лишь через три дня после своего рождения. Этот опыт был перенят и уже через два года, в 2003 году, ученые создают клон особи вола, так же стоящего на грани исчезновения. Вскоре 3 африканских диких кошки были клонированы из замороженных эмбрионов, которые были использованы в качестве ДНК. Несмотря на то, что некоторые эксперты считают, что клонирование спасает особи, стоящие на гране вымирания; некоторые ученые считают, что клонирование несет негативный характер, так как все особи имею генетически идентичный набор хромосом, что в целом играет отрицательную роль, так как для выживания разновидности необходимы разные варианты ДНК.
Некоторые люди так же проявили интерес в том, чтобы их умерших домашних любимцев клонировали, надеясь, что эти клоны будут абсолютно такими же как и их умерший донор. Но как показало клонирование Кошки Сиси, клон не всегда выглядит так же, как и его "оригинал", у которого было взято ДНК [4].
Репродуктивное клонирование - очень неэффективная техника и большинство клонированных животных эмбрионов, не могут развиваться в здоровых особях. Например, Долли была единственным клоном, который был рожден живым из общего количества 277 клонированных эмбрионов. Эта очень низкая эффективность, объединенная беспокойствами по поводу безопасности, представляет серьезное препятствие для применения репродуктивного клонирования. Исследователи выявили некоторые проблемы со здоровьем у овцы и других млекопитающих, которые были клонированы. Это увеличение размера плода при рождение и разнообразные дефекты в жизненных органах, типа печени, мозга и сердца. Другими последствиями являются преждевременное старение и проблемы с иммунной системой.
Другая потенциальная проблема заключается в возрасте хромосомы клонируемой клетки. Все клетки проходят их нормальные стадии деления. Кончик хромосомы, который называется теломером с каждым делением укорачивается. Через какое-то время теломер становится настолько маленьким, что клетка не может больше делится, и в конечном итоге погибает. Это обычный процесс старения, который присущ всем типам клеток. Следовательно, клоны, созданные от клетки, принятой от взрослой особи, могут иметь хромосомы, которые уже короче, чем нормальная, и это может повлиять на быстрое старение клонированной особи. И действительно, Долли, которая была клонирована от клетки шестилетней овцы, имела хромосомы, теломеры которого были короче, чем у овец ее возраста. Долли умерла в возрасте 6 лет, приблизительно половина продолжительности жизни овцы, которая составляет 12 лет [27].
6. Эффективность клонирования животных
Клонирование млекопитающих методом переноса ядер сопровождается патологией в эмбриональный, плодный и неонатальный периоды развития клонов. Возможными причинами аномалий у клонированных животных могут быть ошибки репрограммирования генома, повреждение наследственного материала при культивировании эмбрионов in vitro и сама процедура переноса ядер [9].
Значительная часть выживших клонов обладает рядом нарушений, возникающих из-за несоответствующего эпигенетического репрограммирования генома. Суть проблемы заключается в том, что развитие клонированного животного происходит за счет реализации генетической информации, заключенной в хромосомах донорского ядра из дифференцированной соматической клетки, при этом состояние активности разных генов в соматических и эмбриональных клетках значительно отличается. В соматических клетках в активном состоянии находятся гены, характерные для дифференцированной ткани и ответственные за синтез специфических белков. В то же время для развития эмбриона на ранних стадиях требуется, чтобы синтезировались совершенно другие белки, информация о которых закодирована в генах раннего развития.
В процессе естественного полового размножения зигота образуется в результате слияния мужской и женской половых клеток. В процессе гаметогенеза в половых клетках происходит ремоделирование наследственного материала, т.е. его подготовка к дальнейшим процессам оплодотворения и раннего развития. В момент оплодотворения их гены "молчат", с них не происходит считывания информации. Ядро соматической клетки при его переносе в энуклеированный ооцит не "молчит", в нем активно происходят процессы транскрипции (считывания) [21].
Первые успешные опыты по клонированию доказали, что соматическое ядро в цитоплазме ооцита подвергается репрограммированию - процессу переориентирования донорского генома на синтез белков, соответствующих раннему зародышу. Становятся активными те участки хромосом, которые усиленно работают у раннего зародыша. Во многих случаях процесс репрограммирования генома ядерного трансплантата является неполным, что и приводит к ранней остановке развития эмбрионов.
Существует и другая проблема, связанная с так называемым геномным импринтингом. Явление геномного импринтинга состоит в том, что для нормального развития организма необходимы гены как отцовского, так и материнского происхождения. Известно, что определенные гены так импринтированы в процессе гаметогенеза, что после процесса оплодотворения экспрессируется только отцовский или только материнский аллель. При переносе соматического ядра в энуклеированный ооцит этот механизм может нарушаться, поскольку после их слияния происходит сложная функциональная перестройка всего клеточного генома, в течение которой велика вероятность ошибок.
В данных работах приводится информация о различиях в организации материнского и отцовского хроматина, подчеркивается роль метилирования в подавлении активности генов. Высказывается предположение, что причиной низкой выживаемости животных, полученных в результате переноса ядер, являются генетические нарушения (мутации), аккумулирующиеся в процессе старения клеток организма-донора ядер или во время их культивирования в условиях in vitro.
Высокая частота возникновения аномалий и их межвидовое сходство, а также получение здорового потомства от животных-клонов говорит в пользу эпигенетической природы возникновения таких нарушений, то есть наиболее достоверным объяснением пороков развития является неспособность реконструированных эмбрионов соответствующим образом репрограммировать статус ядра соматической клетки[22].
В работах R. Jaenisсh обсуждается проблема укорачивания концевых участков хромосом (теломер) в клетках клонированных животных. С началом дифференцировки в большинстве клеток происходит необратимое укорачивание концевых участков хромосом, что ставит вопрос о том, наследуют ли клонированные животные укороченные теломеры их генетических родителей и подвержены ли они вследствие этого преждевременному старению. Укорачивание теломер зарегистрировано у первой клонированной овцы Долли, но не отмечено у клонированных в 2000 г. телят. Было установлено, что активность фермента теломеразы, удлиняющего концевые участки хромосом, в ядерных трансплантатах находится на уровне, сходном с контролем. Теломераза полностью восстанавливает длину теломер донорского генома на стадии раннего эмбриона, и, как считает автор статьи, этот фактор не может влиять на выживаемость клонов.
Клонирование методом переноса ядер неодинаково эффективно при использовании в качестве доноров дифференцированных соматических и тотипотентных эмбриональных стволовых клеток. Реконструированные эмбрионы с геномом эмбриональной стволовой клетки, достигшие стадии бластоцисты, развиваются до рождения в 10-20 раз чаще, чем эмбрионы, полученные после переноса ядер соматических клеток. Эти наблюдения дают основания предполагать, что ядру недифференцированной эмбриональной клетки в отличие от дифференцированной требуется лишь незначительное репрограммирование. Это объясняется сходством эпигенетического статуса геномов эмбриональных стволовых клеток и клеток раннего эмбриона. Под эпигенетическим статусом в генетике развития понимается сумма всех взаимодействий генов со средой их функционирования.
Ранее было показано, что родившиеся животные-клоны часто проявляют признаки нарушения дыхания и кровообращения, при рождении имеют повышенный вес тела и плаценты, вследствие чего этой патологии был присвоен термин "синдрома крупного молодняка". Авторами данных статей, однако, не было установлено какой-либо взаимосвязи между изменением активности ряда импринтированных генов у клонов и повышенным весом; аномалии в экспрессии генов носили случайный характер.
Полученные результаты свидетельствовали о значительных вариациях в активности и уровне метилирования импринтированных генов в плацентах и тканях мышат, полученных в результате трансплантации ядер из эмбриональных стволовых клеток. Чтобы выяснить, являются ли эти нарушения результатом изменения импринтинга в донорской популяции ЭС клеток или следствием неправильного репрограммирования донорского генома после пересадки ядер, у нескольких линий ЭС клеток вызывалась направленная дифференцировка добавлением ретиноевой кислоты. В результате выяснилось, что имеются значительные вариации в уровне экспрессии импринтированных генов не только между разными линиями ЭС клеток, но и между различными субклонами одной линии ЭС клеток (т.е. между ЭС клетками, берущими начало от одной-единственной родительской клетки). Эти аномалии возникают в процессе культивирования ЭС клеток в условиях in vitro[27].
Независимо от типа клеток, используемых в качестве доноров ядер, только небольшой процент реконструированных эмбрионов (1-3%) развиваются до рождения, из них менее половины достигают стадии половозрелости. Это поднимает вопрос о том, имеет ли вообще место нормальная регуляция взаимодействия генов у клонированных животных. Рождение здоровых клонов может объясняться толерантностью развития млекопитающих к большей части эпигенетических нарушений, а летальный эффект вызывается кумулятивным действием потерь нормальной регуляции генов во многих локусах.
В заключение авторами статей делается вывод, что, по всей вероятности, клоны всех видов млекопитающих, включая людей, будут обладать эпигенетическими аномалиями и связанным с ними фенотипом. Поскольку эмбриональные стволовые клетки являются потенциальным источником многих типов клеток для использования в целях трансплантационной терапии, очень важно установить, характерно ли состояние эпигенетической нестабильности для ЭС клеток человека, и оценить значение этого явления на перспективы применения ЭС клеток в медицине[13][24].
Выводы
Клонирование – процесс создания генетически сходного организма несексуальным (неполовым) путем. Клонирование использовали много лет для выращивания растений. Животное клонирование было предметом изучения для ученых многие годы, но получало мало внимания до 1997, пока не было клонировано первое млекопитающее - овечка Долли. Ученный Долли и несколько других ученых клонировали различных животных, включая коров и мышей. Недавний успех клонирования привел к жестким дебатам среди ученых, политиков и широкой публики об использование и этике клонирования животных и возможно человека.
За последние 50 лет, ученые провели эксперименты по клонированию в обширном круге животных, использовав много различных методов. В 1979, исследователи произвели первых генетически идентичных мышей, расколов эмбрион мыши в экспериментальной трубе, а затем внедрив получившийся эмбрион в матку взрослой самки мыши. Вскоре после того, как исследователи произвело первых генетически идентичных коров, овцу и цыплят, перемещая ядро клетки, взятой у раннего эмбриона в яйцо, у которого было освобождено ядро.
Главная причина клонирования животных в том, чтобы произвести организмы с определенными качествами, которые необходимы человеку, например овца была выведена чтобы предоставить человеческий инсулин. Если бы ученые полагались только на половое (сексуальное) размножение чтобы вывести этих животных, они бы рисковали тем, что необходимые им качества исчезли, так как половое размножение (сексуальное) переставляет генетический код в блоках. Другими причинами для клонирования могут быть потерянные или умершие домашние животные или животные, которые находятся на грани вымирания. Какими бы не были причины, новые технологии клонирования разожгли много этических спорах среди ученых. Некоторые государства рассмотрели или предписали законодательство, чтобы замедлить, ограничить или запретить эксперименты клонирования. Ясно, что клонирование будет частью нашей жизни в будущем, но будущее этой технологии должно всё же быть определено.
Список использованной литературы
1. С.И.Заир – Бек, И.В.Муштавинский, Развитие критического мышления на уроке, - М.;Просвещение,2004.
2. Газета "Биология" Издательского дома "Первое сентября", № 12/2003.(Л.В. ЯКОВЕНКО Медицина и клонирование)
3. Газета "Биология" Издательского дома "Первое сентября", № 30/2003.(Голем третьего тысячелетия. Религиозные и исторические последствия клонирования)
4.Газета "Первое сентября" Издательского дома "Первое сентября", № 12/2003 ( Василий ЧЕШИРСКИЙ .Самая тонкая цепочка. Что такое клонирование с биологической точки зрения?)
5.Газета "Первое сентября" Издательского дома "Первое сентября", № 15/2003. (А.ВОЛКОВ. Клонирование: мертвый сезон)
6. Газета "Первое сентября" Издательского дома "Первое сентября", № 15/2003.(А.ГРУДИНКИН, Рождены по ошибке. Клонированные организмы чаще всего нежизнеспособны. Почему?) 7. Афонькин С. Ю. Долли бросает вызов, или размышления о клонировании людей. Биология. N 6, 1999.
8. Бутенко Р. Г. Биология клетки и биотехнология. Наука и человечество, 1987.
9. Дейвор Сольтер. Разведение овец путем пересадки клеточных ядер. Биология. N 38, 1997.
10. Кот М. М. Селекция животных. Перспективы развития. Биология в школе. N 2, 1991.
11. Преждевременное старение Долли (обзор журнала "Nature"). Знание-сила. N 9-10, с. 10, 1999.
12. Рувинова Э. И. Еще раз о клонировании. Биология. N7, 1998.
13. Чижиков Максим. Клонирование, сэр! Комсомольская правда. 5 апреля 2000.
14. Чикин Максим. Овечка Долли стареет не по дням, а по часам! Комсомольская правда. 1999.
15. Чойрыш А. И. Правовые и этические проблемы клонирования человека. Государство и право. N 11, 1998, с. 87-93.
16. В.А.Струнников "Клонирование животных: теория и практика", "Природа", №7, 1998 г.
17. Е.В.Мохов. "В ЧЕМ СЕКРЕТЫ ДОЛГОЛЕТИЯ. ПОЧЕМУ МЫ ЖИВЕМ ТАК МАЛО?.."
18. Соровский образовательный журнал, 1999 №4, клонирование животных, Л.И.Корочкин.
19. Журнал "Человек", 1998 №3, Долли – случайность или закономерность? Конюхов Б.В.
20. Журнал "Свет: природа и человек", 1999 №1.
21. Журнал "Студенческий меридиан", 2001 январь.
22. Журнал "Природа", 1998 №7, клонирование животных: теория и практика Струнников В.А.
23. Афонькин С. Ягнята Франкенштейна //Химия и жизнь. - 1999. - № 3.
24. Краснопольская И. Медицина, которой мы не знали // Российская газета. - 2002. - №40 (2908).
25. Кирпанев В.П. Этика клонирования: жизнь или смерть? - Ставрополь, 2004.
26. Дягтерев Н.Д. Клонирование: Правда и вымысел - 128 с. Наука и жизнь: За гранью очевидного
27. Корочкин Л.И. Клонирование Наука сегодня
28. Н. Грин, У. Стаут, Д. Тейлор, Биология, Москва, "Мир", 1993 г.
29. Ф. Киберштерн, Гены и генетика, Москва, "Параграф", 1995 г.
30. Стивен Вир "Клонирование человека аргументы в защиту" //Русская газета
31.Кузина С., Черкасов И: "Клонирование человека: Происки дьявола или победа науки" // Комсомольская правда, 2007 г.
32. Кутковец Т.И., Юдин Б.Г. Уроки незаконченной дискуссии // Человек. 1998
33. Баев А.А. "Геном человека": некоторые этико-правовые проблемы настоящего и будущего//Человек, 1995, №2
34. Дубинин Н.П. Генетика вчера, сегодня, завтра. М., "Советская Россия", 1981

Нравится материал? Поддержи автора!
Ещё документы из категории биология:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ