Низшие хордовые и их филогенетические связи с другими типами животных
1

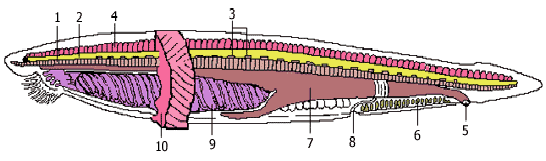

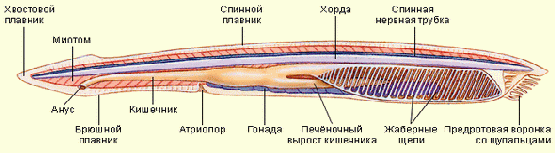
Хордовые и их происхождение
1. ОБЩАЯ ХАРАКТЕРИСТИКА
Хордовые - один из крупнейших типов животного царства, представители которого освоили все среды обитания. В состав этого типа входят три группы (подтипа) организмов: оболочники (аппендикулярии, асцидии, сальпы), бесчерепные (похожие на маленьких рыбок морские существа - ланцетники), позвоночные. Подтипы бесчерепные и оболочники обычно называют низшишими хордовыми, противопоставляя им высших хордовых- позвоночных.
Несмотря на исключительное разнообразие хордовых, все они обладают рядом общих черт строения и развития. Основные из них таковы:
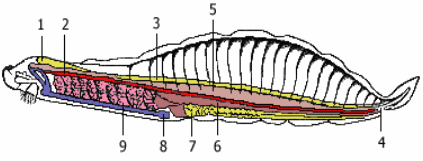
1. У всех хордовых имеется осевой скелет, первоначально возникающий в виде спинной струны, или хорды рис.1. Хорда представляет собой упругий нечленистый тяж, эмбрионально развивающийся путем отшнуровывания его от спинной стенки зародышевой кишки: хорда имеет энтодермальное происхождение. Последующая судьба хорды различна. Пожизненно она сохраняется только у низших хордовых (за исключением асцидии и сальп). У большинства представителей хорда в той или иной мере редуцируется в связи с развитием позвоночного столба. У высших хордовых она является эмбриональным органом и у взрослых животных в той или иной мере вытесняется позвонками, в связи с этим осевой скелет из сплошного нечленистого тяжа становится сегментированным
Рис.1 Строение хордовых на примере ланцетника
2. Над осевым скелетом располагается центральная нервная система, представленная полой трубкой. Полость нервной трубки носит название невроцеля. Трубчатое строение центральной нервной системы характерно практически для всех хордовых. Исключение составляют лишь взрослые оболочники. Почти у всех хордовых передний отдел нервной трубки разрастается и образует головной мозг. Внутренняя полость сохраняется в этом случае в виде желудочков головного мозга. Эмбрионально нервная трубка развивается из спинной части эктодермального зачатка.
3. Передний (глоточный) отдел пищеварительной трубки сообщается с наружной средой двумя рядами отверстий, получивших название висцеральных щелей. У низших форм на их стенках располагаются жабры. Жаберные щели пожизненно сохраняются только у низших водных хордовых. У остальных они появляются лишь как эмбриональные образования, функционирующие на некоторых стадиях развития или не функционирующие вовсе. Наряду с указанными тремя основными особенностями хордовых должны быть упомянуты следующие характерные черты их организации, которые, однако, кроме хордовых имеются и у представителей некоторых других групп.
Хордовые, так же как и иглокожие, имеют вторичный рот. Он образуется эмбрионально путем прорыва стенки гаструлы на конце, противоположном гастропору. На месте же зарастающего гастропора формируется анальное отверстие.
Полость тела у хордовых вторичная (целом). Этот признак сближает хордовых с иглокожими и кольчатыми червями. Метамерное расположение многих органов особенно четко выражено у зародышей и низших хордовых. У высших их представителей в связи с общим усложнением строения метамерия выражена слабо.
Хордовым свойственна двусторонняя (билатеральная) симметрия тела. Как известно, этим признаком кроме хордовых обладают многие группы беспозвоночных животных.
2. ПОДТИП I. БЕСЧЕРЕПНЫЕ (ACRANIA)
Подтип включает один класс Головохордовые (Cephalochordata). В этом классе один отряд - ланцетникообразные (Amphioxiformes) с одним семейством лащетниковые и тремя подсемействами: обыкновенные ланцетники (Branchiostomidae), эпигонихты (Epigonichtys) и амфиоксиды (Amphioxidae).
Тело окаймлено плавниковой складкой: спинной плавник, хвостовой, две складки от предротовой воронки сливаются с подхвостовым плавником. В месте слияния располагается отверстие (атриопор) атриальной полости, биологическое значение которого связано с образом жизни ланцетника.
Тело покрыто однослойным эпителием и кутикулой, под ними располагается слой соединительной ткани (производное мезодермы).
Фотография ланцетника
Скелет. Основу опорной системы составляет хорда, представляющая систему поперечных мышечных пластинок, окруженных соединительнотканной оболочкой. Хорда на переднем конце простирается дальше нервной трубки. К хорде примыкает 50-80 мышечных сегментов (миомер), разделенных перегородками (миосептами). Миомеры состоят из поперечнополосатых мышц. Перегородки связаны с оболочкой хорды и кориумом. Сокращение миомер обеспечивает движение животного.
Центральная нервная система в виде трубки с невроцелем, выраженного головного мозга нет, невроцель в передней части расширен. В стенках передней части нервной трубки обнаружены ядра нейросекреторных клеток, близких к примитивной гипоталомической системе. В стенках нервной трубки имеются светочувствительные органы - глазки Гессе (рис. 2) Периферическая нервная система образована нервами, отходящими от трубки. Работу миохордального комплекса координируют клетки Роде, связанные с нервной трубкой. Органы чувств: обонятельная ямка, органы химического раздражения, механорецепторы (осязательные клетки), осязательные щупальца вокруг ротового отверстия.
Рис. 2. Продольный разрез ланцетника: 1 - хорда, 2 - нервная трубка, 3 - глазки Гесся, 4 - миомеры, 5 - анус, 6 - гонады, 7 - кишечник, 8 - атриапор, 9 - глотка, 10 - мышечный слой.
Кровеносная система ланцетника замкнутая, она примитивна и представлена потоками артериальной и венозной крови Сердца у ланцетника нет. В связи с отсутствием у ланцетника сердца ток крови обусловлен пульсацией брюшной аорты и нижних частей жаберных сосудов ("жаберные сердца"). Кровь бесцветна: она не содержит ни форменных элементов, ни пигмента.
Пищеварение: По способу питания - фильтраторы. С глоткой связаны жаберные щели, ток воды создается реснитчатым эпителием жаберных перегородок и паруса (перегородка вокруг рта). В глотке имеется система желобков с ресничками и железистыми клетками. Пищеварительные ферменты выделяет печеночный вырост. Переваривание пищи происходит и в кишечнике, и в печеночном выросте, характерно и внутриклеточное пищеварение (у позвоночных подобный способ отсутствует).
Дыхание: Специальных органов дыхания нет. Газообмен проходит в стенках глотки, а также по всей поверхности тела.
Выделительная система представлена многочисленными (до 90 пар) нефридиями, расположенными в области глотки.
Половая система, особенности размножения и развития (рис. 3)
Бесчерепные раздельнополы: у каждой особи развиваются яичники либо семенники, внешне похожие: это округлые вздутия на стенке тела, примыкающей к атриальной полости. У каждой особи развивается около 25 пар половых желез. Они не имеют протоков, и зрелые половые клетки выпадают в атриальную полость через разрыв стенки половой железы; подхватываются там током воды и через атриопор выводятся наружу.
Рис. 3 Половая система на вскрытом ланцетнике: 1 - расширение нервной трубки, 2 - спинная аорта, 3 - нервная трубка, 4 - анус, 5 - нотохорд, 6 - кишечник, 7 - гонады, 8 - венозный синус, 9 - жаберные щели.
Выделение зрелых половых продуктов обычно происходит сразу после захода солнца. Подобная синхронность облегчает оплодотворение, которое всегда происходит в воде, вне материнского организма.
Индивидуальное развитие ланцетника типично для хордовых, в том числе и низших позвоночных. Его начальные стадии протекают быстро. Оплодотворенное яйцо (диаметр около 0,1 мм) испытывает полное и почти равномерное дробление, в результате которого образуется шаровидная бластула. Через 5-8 ч после оплодотворения начинается инвагинация: формируется двухслойная гаструла. В период 12-24 ч происходит образование мезодермы (среднего зародышевого листка) и дальнейшая дифференцировка зародыша; формируются нервная трубка, хорда, целомические мешки. Целомические мешки , разрастаясь, подразделяются на лежащий по бокам хорды и нервной трубки толстостенный сомит и ниже расположенную тонкостенную боковую пластинку. Дифференцируясь, стенки сомитов образуют соединительнотканную оболочку хорды и нервной трубки, миомеры и соединительнотканный слой кожи - кутис. Стенки боковых пластинок дают листки брюшины и мускулатуру кишки, а слившиеся полости боковых пластинок превращаются во вторичную полость тела - целом.
На этой стадии в возрасте около суток зародыш разрывает оболочку яйца и выходит в воду в виде личинки длиной около 3 мм. Через 30-36 ч после вылупления на левой стороне переднего конца тела образуется углубление, в которое позже прорывается полость кишечника; так асимметрично закладывается ротовое отверстие. На брюшной стороне тела, позади ротового отверстия возникают жаберные щели, позднее перемещающихся на правую сторону. Затем ротовое отверстие перемещается на брюшную сторону, и личинка становится симметричной. К этому времени прорывается анальное отверстие, а на дне глотки формируется эндостиль, после этого личинка начинает питаться. По бокам тела, над жаберными щелями образуются продольные выросты - метаплевральные складки. Разрастаясь книзу и навстречу друг другу, они смыкаются под брюшком животного, образуя атриальную полость. В виде слепых выростов стенок тела у верхней части глотки возникают нефридии, позже прорывающиеся в атриальную полость. Личиночная стадия продолжается около 3 месяцев
3. ПОДТИП II. ЛИЧИНОЧНО-ХОРДОВЫЕ (UROCHORDATA), ИЛИ
ОБОЛОЧНИКИ (TUNICATA)
Подтип содержит 3 класса: асцидии (Ascidiae), салъпы (Salpae), аппендикулярии (Appendiculariae)
Оболочники. Верхний ряд – асцидии, слева направо: асцидия ментула, колония ботриллуса Шлоссера, клавелина, желудочная циона. Нижний ряд, слева направо: аппендикулярия ойкоплевра, бочёночник долиолетта, колония сальп пегия, атлантическая пиросома
Группа примитивных хордовых животных, которые в личиночной стадии развития имеют все характерные для типа Chordata черты строения, но при переходе во взрослое состояние теряют хорду и испытывают глубокое преобразование центральной нервной системы, которая превращается нз нервной трубки в компактный нервный узел (лишь аппендикулярии сохраняют хорду и нервную трубку всю жизнь!. Упрощение организма с возрастом животных связано с переходом от подвижного существования личинки к неподвижному взрослых особей.
Специфические черты строения: имеется кожно-мускульный мешок (эпителий и слои продольных и кольцевых мышц); кровеносная система незамкнутая, сердце трубчатое, кровообращение маятникообразное; нервная система представлена нервным ганглием, не имеющим внутренней полости, от которого отходят нервные тяжи; выделительная система отсутствует; гермафродиты, оплодотворение во внешней среде. Асцидии и сальпы размножаются и бесполым путем.
Тело оболочников никогда не бывает сегментировано, хотя у некоторых асцидий в нем и заметно подразделение на 2 или 3 отдела. Снаружи тело одето студенистой, кожистой или хрящеватой оболочкой-туникой. Основу ее составляет вещество, крайне близкое к растительной клетчатке (целлюлозе).
Мускулатура. Под наружным эпителием лежит слой соединительной ткани с заключенными в нем мускулами; мускулатура у асцидий состоит из продольных и поперечных мышечных волокон, у сальп образует ряд колец.
Нервная система. Центральная нервная система у взрослых оболочников состоит из одного узла на спинной поверхности с отходящими от него нервами.
Органы чувств мало развиты: встречается глаз в виде пигментного пятна на нервном узле, иногда с преломляющим свет тельцем (у личинок асцидий, у сальп, пирозом), слуховой орган в виде непарного отоциста (у личинок асцидий, у Doliolum), органы осязания в виде выростов на краях вводного и выводного отверстий. Под нервным узлом стенка жаберного мешка вдается, образуя орган, который сравнивают с Hypophysis мозга позвоночных.
Пищеварительная система. Наиболее характерной особенностью кишечного канала является сильное развитие переднего отдела, служащего органом дыхания и принятия пищи. У аппендикулярий стенка этого отдела (жаберного мешка) пронизана лишь двумя отверстиями, которые непосредственно открываются наружу; у асцидий стенка жаберного мешка снабжена многочисленными отверстиями (жаберными щелями), которые открываются в так называемую перибранхиальную или периторакальную полость, окружающую большую часть стенки жаберного мешка и составляющую передний отдел полости клоакальной. Кровеносная с-ма. Сердце лежит на брюшной стороне тела; у аппендикулярий кровеносных сосудов нет, у остальных оболочников от сердца отходит передний и задний сосуд. Замечательная особенность кровообращения О. заключается в том, что сердце некоторое время сокращается в известном направлении, потом сокращения останавливаются и затем начинаются снова, но уже в противоположном направлении; определенного направления движение крови, следовательно, не имеет и в каждом сосуде и в сердце кровь движется то в одном направлении, то в другом.
Половая система и особенности размножения. Все половые особи оболочников - гермафродиты, т. е. они обладают как мужскими, так и женскими половыми железами. Созревание мужских и женских половых продуктов всегда происходит в разное время, и поэтому самооплодотворение невозможно. У асцидий, сальп и пиросом протоки половых желез открываются в клоакальную полость, а у аппендикулярии сперматозоиды выходят в воду через протоки, открывающиеся на спинной стороне тела, тогда как яйца могут выйти наружу только после разрыва стенок тела, что ведет к гибели животного. Оплодотворение у большинства оболочников происходит в клоаке, но имеется и наружное, когда сперматозоид встречается с яйцеклеткой в воде и там ее оплодотворяет. У сальп и пиросом образуется только по одному яйцу, которое оплодотворяется и развивается в теле матери.
Следует подчеркнуть, что приобретение пелагическими туникатами подвижности привело к утрате ими развитой свободноплавающей личинки. У сложных и у большинства одиночных асцидий оплодотворение яиц происходит в клоакальной полости матери, куда с током воды через сифоны проникают сперматозоиды других особей, а оплодотворенные яйца выводятся через анальный сифон. Иногда зародыши и развиваются в клоаке и лишь потом выходят наружу, т.е. имеет место своего рода живорождение.
Сидячим организмам для их успешного размножения необходимо, чтобы яйцеклетки и сперматозоиды у соседних особей созревали одновременно. Эта синхронизация достигается тем, что половые продукты, выведенные наружу первыми половозрелыми особями, с током воды попадают через вводной сифон к соседним животным и в короткий срок стимулируют начало их размножения на значительных территориях. Особую роль при этом играет околонервная железа, сообщающаяся спелостью глотки, воспринимающая соответствующий сигнал из воды. Через нервную систему она ускоряет созревание гонад.
Таблица. Черты сходства и различия личиночнохордовых и бесчерепных
Признак
Бесчерепные
Оболочники
Сегментация тела
Сегментировано
Несегментировано
Опорная система
Хорда
Хорда только на личиночной стадии, искл. аппендикулярии
Питание
Фильтраторы
Фильтраторы
Кровеносная система
Замкнутая, нет сердца
Незамкнутая, сердце трубчатое
Нервная система
В виде трубки с невроцелем, отсутствуют спинномозговые ганглии, выраженного головного мозга нет
Представлена ганглием, мозга нет
Выделительная система
Нефридии
Нет
Размножение
Раздельнополы
Гермофродиты
Оплодотворение
В воде (наружное)
Внетреннее, реже наружное
Развитие
С личинкой
С личинкой
Покров тела
Покрыто однослойным эпитэлием и кутикулой
Покрыто туникой
4. СРАВНЕНИЕ ХОРДОВЫХ С ДРУГИМИ ВТОРИЧНОРОТЫМИ
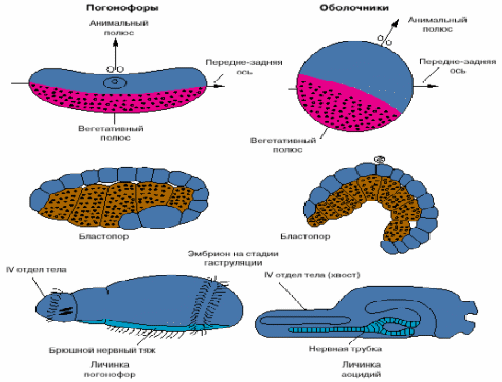
Хотя хордовые - это вторичноротые животные, все же их развитие имеет ряд особенностей. На рис. 4 представлены соответствующие стадии развития одной из групп вторичноротых - погонофор и низших хордовых - асцидий.
Анимальный полюс яйца погонофор, на котором располагается ядро яйцеклетки, соответствует спинной стороне эмбриона, а вегетативный, на котором концентрируется желток, - брюшной. У низших хордовый ситуация обратная: анимальный полюс соответствует брюшной стороне, а вегетативный - спинной. Бластопор (то есть та область эмбриона, где впячивается энтодерма и формируется первичный кишечник) у погонофор соответствует брюшной стороне личинки, а у низших хордовых - спинной. Зачаток нервной системы, закладывающийся по линии замыкания бластопора, у погонофор становится брюшным нервным тяжем, а у хордовых - спинной нервной трубкой (рис. 4).
Рис. 4. Особенности эмбрионального развития погонофор и низших хордовых.
План строения низших хордовых также существенно отличается от планов строения других вторичноротых. Так, например, у всех вторичноротых передние целомы диссимметричны: левый передний целом больше правого, который обычно выполняет функцию околосердечной сумки. У хордовых ситуация обратная: правый передний целом больше левого. Целомы вторичноротых сообщаются с внешней средой отверстиями, открывающимися на спинной стороне и выполняющими осморегуляторные и половые функции. У хордовых эти отверстия располагаются на той стороне тела, которую мы традиционно считаем брюшной.
Кровеносная система у вторичноротых и хордовых устроена по общему плану и состоит из двух главных сосудов: спинного и брюшного. У вторичноротых, как и всех других беспозвоночных, кровь по спинному сосуду течет вперед, а по брюшному назад. У хордовых направление тока крови обратное: по спинному сосуду кровь течет назад, а по брюшному - вперед. Сердце у вторичноротых лежит на спинной стороне, а у хордовых - на брюшной.
Суммируем все эти отличия.
1. Бластопор хордовых соответствует не брюшной, а спинной стороне.
2. Нервная пластинка хордовых, закладывающаяся по месту замыкания бластопора, превращается не в брюшной нервный тяж, а в спинную нервную трубку.
3. Диссимметрия первой пары целомов имеет обратный знак в сравнении с другими вторичноротыми: правый передний целом у хордовых больше левого.
4. Отверстия целомов у хордовых открываются не на спинной, а на брюшной стороне.
5. Кровь у хордовых по брюшной стороне течет вперед, а по спинной - назад, и сердце располагается не на спинной, а на брюшной стороне.
Это значит, что предки хордовых в своем эволюционном развитии испытали инверсию сторон тела, то есть перевернулись и стали передвигаться на морфологически спинной стороне, которая стала функционировать как физиологически брюшная. Морфологически брюшная сторона предков хордовых стала функционировать как физиологически спинная.
Если мы примем этот вывод, все перечисленные выше несоответствия исчезнут: бластопор хордовых станет соответствовать их исходно брюшной стороне, нервная трубка будет располагаться на морфологически брюшной стороне, диссимметрия передних целомов приобретет нормальный для вторичноротых характер, отверстия целомов окажутся на морфологически спинной стороне, и направление тока крови, так же как положение сердца, окажется типичным для вторичноротых. На рис. 5 представители низших хордовых (личинка асцидий и ланцетник) изображены перевернутыми, чтобы продемонстрировать, что при таком положении их системы органов соответствуют таковым других вторичноротых.
Переворот на спинную сторону мог стать тем важнейшим эволюционным событием, которое определило формирование плана строения хордовых животных и его коренное отличие от планов строения вторичноротых и других беспозвоночных.
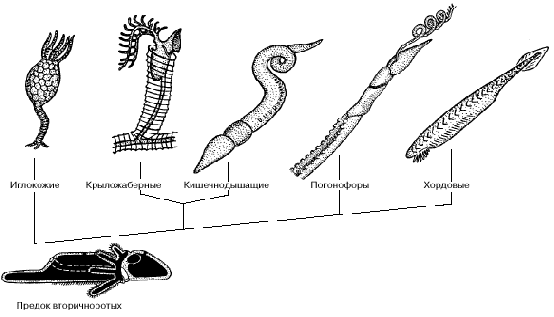
5. ПУТИ ЭВОЛЮЦИИ ВТОРИЧНОРОТЫХ ЖИВОТНЫХ
Общий предок вторичноротых животных, по-видимому, вел донный образ жизни, ползая на брюшной стороне с помощью ресничной полоски, подостланной вентральным нервным тяжем (рис. 6). Он имел три пары целомических мешков и щупальцевый аппарат, с помощью которого собирал мелкие пищевые частицы. В дальнейшем могло произойти обособление самого заднего отдела тела, с помощью которого предки вторичноротых временно зарывались в верхние слои грунта. В эволюции вторичноротых этому отделу принадлежит очень важное место. Предки иглокожих перешли к сидячему образу жизни и стали собирать взвешенные в воде пищевые частицы с помощью щупалец. Радиальная симметрия иглокожих - это наследие сидячего образа жизни, который среди современных форм сохраняют морские лилии, тогда как морские звезды, морские ежи, офиуры и голотурии способны медленно переползать с места на место.
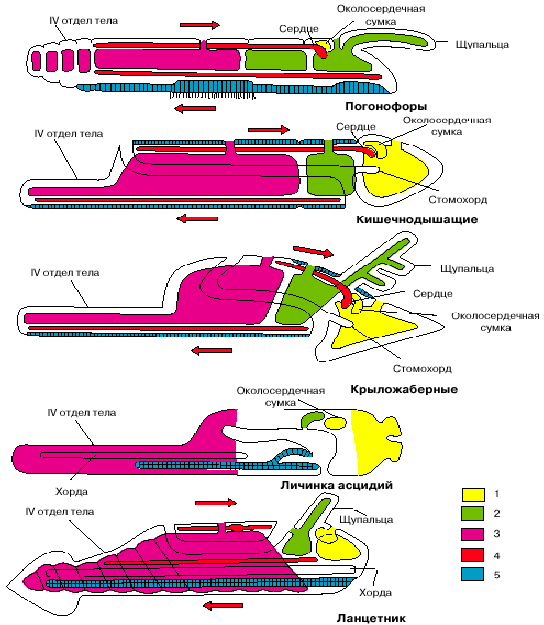
Полухордовые в качестве основного органа движения стали использовать предротовой отдел тела: хобот кишечнодышащих или головной щит крыложаберных. Обычно полухордовых считают близкими предками хордовых, о чем говорит и само название этой группы.
Рис. 5. Сравнение планов строения различных групп вторичноротых (личинка асцидий и ланцетник изображены в перевернутом положении). 1 - целомы первой пары, 2 - целомы второй пары, 3 - целомы третьей пары, 4 - кровеносная система, 5 - нервная система. Стрелки показывают направление тока крови.
В действительности эта группа не слишком близка к хордовым, в частности потому, что имеет два нервных тяжа, только один из которых может быть гомологизирован с нервной трубкой хордовых. У них имеется слепой вырост кишечника - стомохорд, который сформировался как опорная структура хобота и не имеет прямого отношения к хорде хордовых животных.
Современные погонофоры - сидячие организмы, обитающие внутри выделяемых ими трубок. Во взрослом состоянии они лишены кишечника и питаются за счет симбиотических хемосинтезирующих бактерий, окисляющих сероводород или метан. Их предки, вероятно, питались этими бактериями, собирая их из придонного слоя воды с помощью щупалец.
Рис. 6. Филогенетические отношения между вторичноротыми животными.
Впоследствии они стали культивировать бактерий в клетках кишечника. Личинки одной из групп современных погонофор - вестиментифер - на ранних стадиях развития собирают бактерий с поверхности грунта, а затем начинают их культивировать внутри видоизмененного кишечника, после чего рот и анус у них редуцируются. Особенности эмбрионального развития и строения нервной системы позволяют рассматривать погонофор как самую близкую к хордовым группу вторичноротых.
Предки хордовых также первоначально собирали пищевые частицы с помощью околоротовых щупалец, которые до сих пор сохранились у ланцетников. Одной из особенностей эволюции хордовых явилось использование самого заднего, четвертого отдела тела как органа активного движения. Этот отдел имеется у всех вторичноротых. Иглокожие и крыложаберные используют его как прикрепительный стебелек. У кишечнодышащих этот отдел функционирует только у молодых особей: с его помощью они закапываются в толщу грунта. У погонофор этот отдел активно функционирует всю жизнь: он вооружен щетинками, торчит из заднего конца трубки и благодаря его движениям погонофоры заглубляют задний конец трубки в толщу грунта по мере своего роста. Хордовые - единственная группа вторичноротых, использующая этот отдел как орган активного плавания. Именно в связи с активной двигательной функцией этого органа в нем происходит полная сегментация мускулатуры и развивается хорда (у личинок оболочников хорда локализована именно в хвосте. Другой важный момент эволюции хордовых - это инверсия сторон тела, то есть произошедший в филогенетическом развитии хордовых переворот с морфологически брюшной на морфологически спинную сторону. Такой переворот мог совершиться при переходе от улавливания с помощью щупальцевого аппарата взвешенных в толще воды пищевых частиц, как это делают, например, современные крыложаберные, к их сбору из поверхностного слоя грунта, как это делают современные ланцетники.
Заметим, что между личинками оболочников, с одной стороны, и бесчерепными и позвоночными, с другой, есть одно важное различие. Если внимательно посмотреть на две нижние фигуры на рис. 5, изображающие личинку асцидии и ланцетника, то нетрудно заметить, что у первых рот располагается на той же стороне, что и нервная трубка (последняя даже открывается в глотку особым отверстием), то есть на исходно брюшной стороне, а у ланцетника (так же, как у позвоночных) - на противоположной стороне тела. Положение рта у личинок асцидий - первичное, оно совпадает с тем, что имеет место у других вторичноротых (см. рис. 4). Таким образом, личинки асцидий по своей организации более примитивны, чем остальные хордовые, и отражают организацию подвижных предков хордовых до их переворота.
У бесчерепных и позвоночных рот располагается на противоположной стороне, гомологичной спинной стороне тела прочих вторичноротых (см. рис. 5). Такое положение рта не может быть первичным и возникло уже после переворота. Удивительно, но в индивидуальном развитии бесчерепных сохранились следы постепенного перемещения рта со спинной стороны на брюшную по левой стороне тела. А один из представителей бесчерепных (Assymetron) так и застыл на стадии, когда рот остался на левой стороне тела. Все это отголоски постепенного перемещения рта со старой морфологически брюшной стороны на новую физиологически брюшную сторону.
6. ОСНОВНЫЕ ГИПОТЕЗЫ ПРОИСХОЖДЕНИЯ ХОРДОВЫХ
Одна из первых концепций, связывающих план строения хордовых с планами строения других групп беспозвоночных, была разработана выдающимся французским сравнительным анатомом Жоффруа Сент-Илером еще в первой половине XIX века. Он считал, что членистые животные (кольчатые черви и членистоногие) могут рассматриваться как перевернутые позвоночные. "Взгляните на рака, опрокинутого на спину, говорил он, - и вы увидите, что различные системы его расположены совершенно так же, как у высших позвоночных животных". Именно эта концепция была в центре публичного спора между Жоффруа Сент-Илером и Жоржем Кювье, который состоялся в Париже в 1830 году в дни июльской революции, свергнувшей Бурбонов. Как известно, в том знаменитом споре победил Жорж Кювье. Однако к идее сравнения хордовых (и позвоночных) и перевернутых кольчатых червей или членистоногих зоологи возвращались неоднократно на протяжении всего XIX века.
В начале нынешнего столетия были разработаны гипотезы происхождения хордовых от кишечнодышащих - обитающих в толще грунта морских червеобразных организмов. У этих животных (как и у хордовых) имеются жаберные щели, а в передней части тела - опорный вырост кишечника (стомохорд), который сравнивали с хордой. Однако кишечнодышащие резко отличались от хордовых обратным направлением тока крови, расположением сердца, строением нервной системы и другими важными особенностями организации. Делались попытки выводить хордовых от иглокожих, немертин, моллюсков и других групп беспозвоночных, однако ни одна из них не имела успеха.
Своеобразной реакцией на неудачи зоологов решить проблему происхождения хордовых традиционными способами явилась разработка так называемых неотенических гипотез, выводящих хордовых от личинок асцидий. Взрослые асцидии - прикрепленные морские животные, фильтрующие воду через объемистый жаберный мешок и улавливающие взвешенные в воде мельчайшие органические частицы. Они не имеют ни хорды, ни нервной трубки и совсем не похожи на хордовых животных. Но, как давно известно (а впервые это показал еще в конце прошлого века выдающийся русский биолог А.О. Ковалевский), личинки асцидий имеют и хорду, и нервную трубку, которые утрачиваются при метаморфозе. На основании этого многие биологи (Берилл, Уитер, Боун и др.) предполагают, что хордовые произошли от ставших половозрелыми личинок асцидий. Правда, в этом случае остается непонятным, откуда взялись сами асцидии. Да и трудно представить, чтобы хорда и нервная трубка возникли у живущих 1 - 2 дня и не питающихся личинок. Более вероятно, что эти особенности - наследие свободноживущих предков асцидий, об организации которых неотенические гипотезы ничего не говорят. Ни одна из рассмотренных гипотез в настоящее время не может считаться истинной, но в каждой из них содержатся отдельные элементы, которые могут оказаться полезными при реконструкции путей происхождения хордовых.
ЗАКЛЮЧЕНИЕ
Современные представления о происхождении хордовых объединяют положения, существовавшие ранее в рамках альтернативных гипотез происхождения хордовых: перевернутость хордовых, их родство с вторичноротыми и факт большей примитивности личинок оболочников по сравнению с другими хордовыми. В отличие от ранее существовавших точек зрения хордовые рассматриваются не как перевернутые кольчатые черви и членистоногие, но как перевернутые вторичноротые. Что же касается личинок асцидий, то они не являются предками остальных хордовых, как это принимается в гипотезах неотенического происхождения хордовых. Организация личинок асцидий рекапитулирует (то есть повторяет в сжатой, видоизмененной форме) в индивидуальном развитии определенный филогенетический этап в историческом развитии хордовых, а именно строение предков хордовых до их переворота.
Дискуссия о происхождении хордовых не закончена. Развитие биологической науки приносит новые доводы в пользу той или иной точки зрения. Иногда эти доводы приходят с совсем неожиданной стороны. Так, относительно недавно, в 1993 году, в английском журнале "Nature" появилась статья биологов Слэка, Холланда и Грэма, анализирующая многочисленные публикации, в которых были выявлены зоны экспрессии одной интересной группы генов (так называемой Hox-группы) в развитии различных беспозвоночных и позвоночных животных. В этих публикациях было показано, что у беспозвоночных эти гены маркируют брюшную сторону, а у позвоночных их экспрессия наблюдается на спинной стороне. Этот факт хорошо согласуется с идеей перевернутости хордовых, принимаемой современной сравнительной анатомией.
хордовые вторичноротые происхождение эволюция
ЛИТЕРАТУРА
1. Иванова-Казас О.М. Очерки по филогении низших хордовых. СПб.: Изд-во СПб ун-та, 1995. С. 1 - 160.
2. Малахов В.В. Проблема основного плана строения в различных группах вторичноротых животных. // Журнал общей биологии. 1977. Т. 38. N 4. С. 485 - 499.
3. Малахов В.В. Новый взгляд на происхождение хордовых. // Природа. 1982. N 5. С. 12 - 19.
4. В.М. Константинов, С.П. Наумов, С.П. Шаталова. Зоология позвоночных. М: Академия, 2000. 496с.
5. Наумов М.П., Каратышев Н.Н. Зоология позвоночных. – Ч.1. – Низшие хородовые, бесчелюстные, рыбы, земноводные: Высш. Школа 1979. – 333с.

Нравится материал? Поддержи автора!
Ещё документы из категории биология:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ