Розвиток хрящової тканини в онтогенезі безхвостих
 Зміст
Зміст
Вступ 2
Розділ 1. Загальна характеристика хрящової тканини 4
Розділ 2. Порівняння будови та розвитку хрящової тканини безхвостих та ссавців 9
Висновок 19
Список літератури 20
Вступ
З курсу загальної ембріології відомо, що в зиготі запрограмована можливість формування цілого організму. Першими розблоковуються гени, що зумовлюють здатність клітини до проліферації і регулюють загальний метаболізм клітин. В міру того, як продовжується дроблення і зародок вступає в стадію гаструляції, починають активізуватися перші тканино специфічні гени. В різних клітинах і в різні фази розвитку одні гени можуть функціонувати, а інші — бути заблокованими. Тому виникають різні ферментативні системи і різні типи обміну речовин. Відбувається диференціювання клітин, утворюються різні тканинні структури.
Диференціюванням називається процес, в результаті якого клітина стає спеціалізованою. В основі диференціації знаходиться процес детермінації — визначення подальшого напряму в розвитку клітин на генетичній основі — дії окремих компонентів генома. Диференціацію можна розглядати з різних позицій морфологічної, функціональної, молекулярної тощо. Основна теза молекулярної диференціації полягає у тому, що клітинна диференціація ґрунтується на синтезі специфічних білків, тобто клітини, що диференційовані в різних напрямках, різняться між собою за одним специфічним білком — послідовністю амінокислот білкової молекули. На тканинному рівні диференціація також проявляється у вигляді характерних морфологічних і функціональних змін, що відбуваються в певних групах клітин, частинах організму і в певний час. Процес, у результаті якого окремі тканини в ході диференціації набувають характерного для них вигляду, називають гістогенезом. Він відбувається в ембріональному періоді онтогенезу після утворення зародкових листків.
Для земноводних характерна відносна крихкість кісткової тканини, тому можна припустити, що окостенінні деяких кісток проходить не повністю, і деяка частина кісток мають у своєму складі більшість клітин хрящової тканини. Хрящова тканина належить до сполучної тканини і виконує функцію опорну та формоутворюючу.
Мета роботи – розглянути та охарактеризувати особливості розвитку хрящової тканини у безхвостих земноводних.
Для досягнення поставленої мети нам потрібно вирішити деякі завдання:
дати загальну характеристику хрящової тканини;
проаналізувати спільні та відмінні особливості будови хрящової тканини у безхвостих та ссавців.
Розділ 1. Загальна характеристика хрящової тканини
Являє собою спеціалізований різновид сполучної тканини, яку відносять до групи тканин з опорною та формоутворюючою функцією. Основні компоненти хрящової тканини — міжклітинний матрикс та клітини — хондроцити. До матриксу входять волокна та основна міжклітинна речовина. Залежно від переважання колагенових чи еластичних волокон і розвитку основної речовини розрізняють гіаліновий, еластичний і волокнистий хрящі. Матрикс і хондроцити являють собою єдине ціле в будові та функціональному відношенні — хондроцити продукують матрикс, матрикс забезпечує підтримку фенотипу хондроцита.
До складу хрящового матриксу входить до 75% тканинної рідини, яка міститься у гелеподібній структурі і є важливим компонентом в пружинно-еластичних властивостях хряща і, в цілому, його будові та функції. Склад колагену в матриксі хрящової тканини становить від 50% до 70% сухої маси і залежить від різновиду хряща. Колагенові (хондринові) волокна складаються з колагену другого типу, тонкі мають від 10 до 100 нм в діаметрі.
Органічними компонентами основної міжклітинної речовини (хондромукоїд) є білки, ліпіди, сульфатовані глікозаміноглікани. Останні з'єднуються з не-колагеновими білками і утворюють протеоглікани. У складі протеогліканів хряща знаходяться гігантські макромолекулярні комплекси, побудовані з гіалуронової кислоти. До неї нековалентними зв'язками приєднуються поліпептидні ланцюги; з серединними амінокислотними залишками поліпептидних ланцюгів зв'язані полісахаридні ланцюги сульфатованих глікозаміногліканів хондроітин-, кератан-, дерматансульфату, а також молекул олігосахаридів. Молекула протеоглікана нагадує гілочку ялини, ступінь її гідратації впливає на тургор хряща.
Серед клітин хряща розрізняють хондробласти та хондроцити. Перші мало-диференційовані клітини, мітотично активні, походять від стовбурових клітин мезенхіми, в більшій мірі ніж хондроцити мають цитоплазматичні вирости, в їх цитоплазмі відсутні ліпіди та глікоген. У цитоплазмі помітна базофілія, розвинені комплекс Гольджі, гранулярний ретикулум, РНК, мітохондрії; ці клітини здатні продукувати специфічні для хряща компоненти матриксу. За своєю топографією вони тісно пов'язані з охрястям і зберігають овальну форму.
Хондроцити значно більші від хондробластів, округлої або полігональної форми, займають більш центральне положення в товщі хряща. Розміщені в порожнинах (лакунах) матриксу ізольовано або групами з двох-шести клітин. Останні називаються ізогенними групами у результаті розмноження амітозом однієї клітини. За ультраструктурними та функціональними характеристиками виділяють такі типи хондроцитів. До першого типу відносять малодиференційовані мало-чисельні клітини первинної хрящової тканини. Вони мають нерівні відростки цитоплазми, велике ядро, слабо розвинуту ендоплазматичну сітку й добре розвинений комплекс Гольджі, вільні рибосоми та мітохондрії. Клітини діляться мітозом, беруть участь у фізіологічній регенерації при заміні популяції хондроцитів суглобового хряща. Хондроцити другого типу становлять основну масу хондроцитів діаметром до 20 мкм і характерні для кожного різновиду гіалінового хряща, мають низьке ядерно-цитоплазматичне співвідношення, підвищений вміст РНК та гранулярної ендоплазматичної сітки й комплексу Гольджі, які забезпечують утворення та виділення протеогліканів й глікозаміногліканів. Для ходро-цитів другого та третього типів характерна відсутність мітозів, вони репродукуються шляхом амітозу, утворюють ізогенні групи. У хондроцитів третього типу найнижчий показник ядерно-цитоплазматичного співвідношення, синтез протеогліканів та глікозаміногліканів знижений.
Гістогенез хрящової тканини. Розвиток хряща відбувається за рахунок мезенхіми, її клітини інтенсивно розмножуються, збільшуються в розмірі, ущільнюються і перетворюються в острівці передхрящової тканини; в останній відсутня міжклітинна речовина, її опорна функція визначається внутрішньою напругою, зумовленою інтенсивним розмноженням клітин з великим вмістом у них води. В подальшому ці клітини перетворюються у хондробласти, які здатні продукувати міжклітинну речовину з диференціюванням в ній колагенових волокон. Утворена міжклітинна речовина набуває оксифілії. Подальший стан гістогенезу характеризується диференціацією хондробластів, в яких добре розвинені органели — цистерни гранулярної ендоплазматичної сітки, комплекс Гольджі, мітохондрії. Створюються умови для збільшення маси хрящового зародка, що відбувається як за рахунок збільшення кількості міжклітинної речовини, так і розмноження хондробластів, з'являються базофільні ділянки хряща. В міру збагачення міжклітинної речовини клітини хрящової тканини ізолюються в окремих лакунах (порожнинах) і перетворюються у хондроцити. Подальший ріст хрящової тканини забезпечується поділом хондроцитів та утворенням між дочірніми клітинами міжклітинної речовини. Дочірні клітини, що відокремлені одна від другої тонкими перегородами основної речовини утворюють характерні для зрілого хряща ізогенні групи клітин (від isos — рівний, однаковий, genesis — походжен-ня). На пізніших етапах розвитку хряща утворення міжклітинної речовини сповільнюється, інтенсивність розмноження хондроцитів знижується.
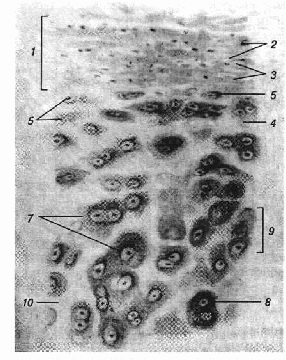
Хрящова тканина (рис. 1), за винятком суглобів оточена сполучною тканиною — охрястям, яке містить нерви та кровоносні судини. Останні забезпечують живлення хрящової тканини та її регенерацію. Розвиток охрястя відбувається в зв'язку з розвитком і ростом хряща.
Рис. 1. Хрящова тканина:
А — еластичний хрящ; Б — волокнистий хрящ
Клітини мезенхіми, які оточують хрящ, що розвивається,
Клітини мезенхіми, які оточують хрящ, що розвивається, залишаються щільно упакованими, із них утворюється порівняно щільна оболонка — охрястя (перихондр). Клітини його внутрішнього шару залишаються малодифереційованими, вони утворюють так званий хондрогенний шар охрястя і здатні проліферувати та диференціюватися в хрящові, утворюючи зовнішню частину хрящової тканини у доповнення до існуючої. Клітини мезенхіми зовнішнього шару диференціюються у фібробласти, які утворюють колаген.
Таким чином, вся структура цілком стає обмеженою волокнистою тканиною, що називається волокнистим шаром охрястя.
Ріст хряща відбувається двома шляхами: амітотичним поділом хрящових клітин і утворенням матриксу — інтерстиціальний ріст — зсередини; накладанням нових шарів хрящової тканини, що утворюються охрястям — наростання зовні — апозиція. Цим пояснюється, чому у малодиференційованого хряща міжклітинна речовина фарбується по-різному, молодші його частини оксифільні, глибші частини міжклітинної речовини мають спорідненість з базофільними фарбниками.
Гіаліновий хрящ. В організмі тварин він найбільш поширений; з нього побудований скелет зародка, в дорослих тварин він входить до складу реберних хрящів у місцях їх з'єднання з грудиною та хребцями, частина гортані містить гіалінові хрящі, вони є в повітроносних шляхах, покривають поверхню епіфізів трубчастих кісток. Своєрідної будови хрящ зовнішнього слухового проходу та слухових труб, в своєму складі вони мають різні види хряща: гіалінового, еластичного та волокнистого.
Зовні гіаліновий хрящ покритий охрястям; на межі з хрящовою тканиною і охрястям знаходяться перехідні форми клітин. Ближче до охрястя вони подібні до клітин останнього, тобто мають веретеноподібну форму, їх довга вісь спрямована вздовж поверхні хряща, у глибоких зонах хрящової тканини клітини поступово збільшуються у розмірі, округлюються. їх поверхня має мікроворсинки,які добре помітні під електронним мікроскопом, ядро хондроцитів шароподібне, бідне на хроматин. Цитоплазма містить глікоген та воду, остання визначає їх високий тургор. У зв'язку з тим, що синтетичні та секреторні процеси в глибоких частинах хряща слабнуть, вони після поділу не розходяться, лежать більше компактно, тому утворюються так звані ізогенні групи (рис. 2).
Клітини обмежені капсулою із колагенових волокон, занурених в аморфну речовину, причому навколо молодих хондроцитів міжклітинна речовина оксифільна. Більш диференційовані хондроцити й ізогенні групи, крім оксифільного перицелюляр-ного шару, мають базофільну зону міжклітинної речовини. Це пояснюється нерівномірним розподілом хімічних компонентів міжклітинної речовини — білків та глікозаміноглі-канів. Клітинні території розділені ок-сифільними або слабобазофільними ділянками — хрящовими балками.
Міжклітинна речовина хряща не має судин, її живлення відбувається шляхом дифузії.
Еластичний хрящ входить до складу вушної мушлі, рожкоподібних та клиноподібних хрящів гортані. На відміну від гіалінового еластичний хрящ характеризується тим, що в його міжклітинній речовині, крім колагенових фібрил, міститься сітка еластичних волокон, здатних витримувати згинання.
Волокнистий хрящ зустрічається у міжхребцевих дисках, круглій зв'язці стегна, а також у місцях прикріплення сухожилків до кісток. Міжклітинна речовина його утворена пучками колагенових волокон з щільної волокнистої сполучної тканини, здатних витримувати значне механічне навантаження. Його клітини округлі з світлою цитоплазмою.
Рис. 2. Гіаліновий хрящ:
1 — охрястя; 2 — колагенові волокна охрястя; 3 — ядра клітин охрястя; 4 — міжклітинна речовина хряща; 5-6 — хрящові клітини; 7 — ізогенні групи; 8 — базофільні зони; 9— клітинні території; 10 — інтертерриторіальна міжклітинна речовина
Розділ 2. Порівняння будови та розвитку хрящової тканини безхвостих та ссавців
Об'єктами дослідження були узяті представники нижчих (безхвості амфібії) і вищих (ссавці) наземних хребетних з тією метою, щоб зіставити стан хрящового покриву в синовіальних суглобах різної морфологічної зрілості. Подібні порівняння можуть допомогти глибше зрозуміти механізми структурного формування суглобного хряща і з'ясувати спірними питання, що залишаються до цих пір, про джерела його самозаповнення і трофіки.
Послідовність формування суглобного хряща й участь у цьому процесі хондроцитів вивчали на хрящовій і кістковій стадіях розвитку кістяка кінцівок (грудних і тазових). Досліджено матеріал від озерної і трав'яної жаб, білого пацюка, морської свинки, домашнього бика. Особливості метаболізму і специфічного біосинтезу в хондроцитах вивчали також у досвідах на білих пацюках і морських свинках, яким уводилися інтраперитонеально в індикаторній дозі радіоактивні попередники біосинтезу ДНК (Н-тимидин), колагену (3Н-гліцин), сульфатованих глюкозаміногліканів (35SО4-сульфат), глікогену (14С-глюкоза). Обробку матеріалу, виготовлення гістологічних зрізів і гістоавтографів проводили загальноприйнятими методами. Змонтовані на предметних стеклах зрізи кістково-хрящових ділянок суглобних кінців кісток забарвлювали гематоксілін-еозином, пікрофуксіном, гематоксилін-тіонін-еозином (Мажуга, Вечерская, 1974). На ультратонких зрізах суглобного хряща за допомогою електронного мікроскопа досліджували стан ядра, гранулярного й агранулярного ендоплазматичного ретикулуму, комплексу Гольджи, мітохондрій, зміст продуктів внутрішньоклітинного вуглеводного, білкового, ліпідного біосинтезів у хондроцитах. Ідентифікацію складу вмісту кліток і міжклітинного хрящового матриксу проводили за допомогою відповідних гістохімічних реакцій і по включенню у визначені субстрати радіоактивних маркерів. Кровоносну систему в компонентах суглоба досліджували методом просвітління після ін'єкції судин чорною тушшю (Мажуга, 1966).
У дорослих особин безхвостих земноводних у деяких суглобах кінцівок (наприклад, у колінному) кінці довгих трубчастих, які зчленовуються, кістки (епіфізи) протягом всього життя зберігають хрящовий стан. У той же час в інших кістякових ланках (наприклад, у лопатко-плечевому суглобі) хрящові епіфізи в основній своїй масі заміщаються кісткою. Тільки вузький суглобний край залишається хрящовим, за аналогією його можна порівняти із суглобним хрящем ссавців. У першому випадку можна говорити лише про суглобні поверхні на сполучених хрящових епіфізах. В другому випадку маємо справу зі збереженої з боку суглоба і периферичною частиною хрящового епіфіза, що перейшла в дефінітивне, стан, що прийняв форму і функцію самостійного компонента - суглобного хряща. І хоча хрящовий суглобної край продовжує залишатися частиною вихідного цілого, усе-таки виникає питання: чи змінилися джерела його трофіки? Іншими словами: чи існують розходження в харчуванні цілого хрящового епіфіза і тієї його периферичної частини, що залишається хрящовий після завершення синостозу?
У ссавців, як відомо, внутрішній кістяк у ранньому онтогенезі також проходить хрящову стадію розвитку. У цей період зчленування утворяться, як і в земноводних, шляхом прямого контакту суглобних кінців хрящових закладок кістяка. Диференціювання суглобного хряща відбувається значно пізніше в процесі заміщення провізорних закладок кістяка кістою. При цьому як у закладках коротких кіст, так і в епіфізах довгих кіст процес заміщення кістою не поширюється на саму периферичну частину суглобного кінця; вона зберігається протягом всього життя індивіда у вигляді хрящового покриття - суглобного хряща. Поступово він набуває своєї особливої внутрішню цитоархітектоніку. При досить обмеженій загальній масі (товщина суглобного хряща виміряється в міліметрах) на гістологічному зрізі суглобного хряща чітко виступає зональний поліморфізм хондроцитів, що на різній глибині залягання відрізняються між собою по обсязі, формі, репродуктивними властивостями, щільністю розподілу в матриксе, ознаками ультраструктурної організації, особливостями метаболізму й ін. Загальна картина цитоархітектоніки дозволяє виділити в суглобному хрящі три структурні зони: поверхневу, звернену до синовіальної порожнини суглоба; середню або проміжну зону і глибоку, звернену до кістки зону. Оскільки вихідними для структурної самопідтримки суглобного хряща є клітини його проміжної зони, доцільно почати опис з неї.
Проміжна зона. По обсязі, займаному в загальній товщині зрілого суглобного хряща, на частку середньої (проміжної) зони приходиться приблизно одна третина (чіткої видимої границі між зонами немає). Хондроцити тут розміщені невеликими ізогенними групами, розміри яких по кількості клітин, що утримуються, помітно зменшуються до поверхневої зони. Ізогенні гнізда являють собою щільно упаковані стовпчики хондроцитів, орієнтовані строго вертикально до суглобної поверхні. Уже саме явище ізогенії свідчить про здатності кліток до саморепродукції, що зберігається в проміжній зоні протягом усього життя. Тому в різні вікові періоди серед кліток ізогенних груп зустрічаються, хоча і з неоднаковою частотою, фігури мітотичного поділу, а на гістоавтографах з 3Н-тимидином - включення цього маркера в ядро хондроцитів, що перебувають у репродуктивному циклі. Трансмісійною електронною мікроскопією в хондроцитах проміжної зони виявляється система добре розвитих цитоплазматичних органоїдів: гранулярний ендоплазматичний ретикулум, апарат Гольджи, мітохондрії, а також різної величини і форми вакуолі, наповнені електроннопрозорими субстанціями і ліпідами. Стан ядра і цитоплазми в клітках проміжної зони, так само як і асиміляція ними радіоактивних попередників нуклеїнового, протеїнового і полісахаридного обмінів виразно відбивають активний стан метаболізму і біосинтезу, спрямованих на задоволення потреб саморепродукції кліток і поповнення компонентів матриксу. Переважними структурами матриксу тут є колагенові волокна, доповнені макромолекулярними агрегатами протеогліканів, глікопротеїдів і хондроітинсульфатів. Описуваний стан є характерним, але не для всієї популяції хондроцитів одноманітним. Навіть у межах окремих ізогенних груп у зрілому хрящі тільки одиниці здатні асимілювати 3Н-тимидин і, приблизно, на 600- 820 хондроцитів можна зустріти одну фігуру мітозу. Для більшості кліток характерне включення екзогенних 35SО4-сульфату, 3Н-глицина,хоча мітки включень цих ізотопів розподіляються серед окремих хондроцитів з неоднаковою інтенсивністю в те саме час на різних територіях проміжної зони. Таке явище обумовлене тим, що процеси біосинтезу розподілені між клітками асинхронно, і в різних субпопуляціях хондроцитів суглобного хряща структурний стан, характер і інтенсивність специфічного біосинтезу послідовно змінюються відповідно їх диференціюванню (дозріванню) і спрямованості однієї убік глибокої зони, інших - поверхневої зони. Тобто гетероморфність і функціональна неоднорідність хондроцитів проміжної зони сполучені з їхньою роллю камбію для всієї структури суглобного хряща. Саме клітки проміжної зони забезпечують структурна самопідтримка суглобного хряща і заповнення його природного збитку. Можливості фізіологічної регенерації всієї тихорєцької системи суглобного покриву закладені в механізмах її внутрішнього (інтерстиціального) росту (Мажуга, 1994).
Поверхнева зона. Усупереч поширеній думці, начебто поверхня нормального суглобного хряща не містить загиблих клітин (Модяєв, 1983; Павлова й ін., 1988), у дійсності це не так. З численних спостережень за станом суглобного хряща в представників різних хребетних з очевидністю випливає, що вся послідовність змін у клітках і матриксі поверхневої зони відбувається по шляху поступового вгасання, деградації і неминучому збиткові їхній з боку суглобної поверхні. Установлено, що матрикс суглобного хряща на 50-70% складається з колагену, має великий запас міцності, повний обмін колагену в ньому може розтягуватися до 120-160 років у людини (Maroudas, 1980), однак, в умовах постійних ковзань у суглобі природне зношування хряща з боку суглобної поверхні відбувається протягом всього індивіда життя. Якщо ж звернутися безпосередньо до гістологічних зрізів суглобного хряща, то на них можна побачити, що клітки в поверхневій зоні не мають ознак репродукції, ближче до суглобної поверхні вони подрібнюються, уплощаються, ядро їхній переходить у стан одночасної прогресуючої редукції цитоплазматичних органоїдів і самої цитоплазми. У самого краю суглобної поверхні видні лише зморщені залишки колишніх хондроцитів. Регресивні зміни відбуваються також у матриксі через послідовно прогресуюче вгасання біосинтезу в хондроцитах його компонентів. Дотепер було прийнято вважати (Stockwell, 1983; Modіs et al., 1986), що спеціалізація хондроцитів на біосинтезі колагену 1-го типу характеризує їхня незрілість і можливість подальшої хондрогенної диференціювання. Це, однак, не можна поширити на хондроцити поверхневої зони суглобного хряща. Здатність їх до синтезу начебто б не властивих зрілому хрящеві субстанцій, зв'язана, треба думати, із прямим впливом на метаболізм у хондроцитах синовіального середовища суглоба. У поверхневі шари суглобного хряща із синовіальної рідини силами компресії щось нагнітається (Дідух і ін., 1992). Цим "щось" практично може бути тільки гіалуронова кислота, і її присутність у поверхневій зоні хряща постійно підтримує особливий метаболічний фон, що виключає можливість прогресивної диференціювання хондроцитів. Вони перебувають тут як би в незрілому стані і ближче до суглобної поверхні, де в матриксе хряща концентрація гіалуронату особливо висока, хондроцити дегенерують і гинуть. Зрозуміло, з руйнуванням хондроцитів і їхньою елімінацією одночасно губляться самі поверхневі шари хрящового матриксу.
Глибока зона. У той період онтогенезу, коли ріст кістякової ланки ще продовжується, з боку глибокої зони суглобного хряща відбувається типовий процес енхондрального заміщення, ідентичний тому, що відбувається в епіфізарній хрящовій пластинці (докладно описано раніше: Mazhuga, 1984; Мажуга, 1992). Проліферація кліток у проміжній зоні в цей період спрямована переважно на поповнення глибокої зони, чим компенсується збиток при руйнуванні гіпертрофованих (набряклих) хондроцитів. В міру резорбції самих нижніх шарів хондроцитів фронт заміщення насувається на чергові їхні ешелони, що досягають до цього часу гіпертрофованого стану. Територія зруйнованих кліток використовується в процесі енхондрального заміщення для адекватного приросту кісткового епіфіза. Тобто, глибока зона суглобного хряща на цьому етапі онтогенезу бере участь у загальному механізмі апозиційного росту кістки.
З завершенням росту кісткової ланки енхондральне заміщення цілком блокується: у паросткових хрящах шляхом синостоза метафіза з епіфізом, у суглобному хрящі - шляхом формування під ним суцільної кісткової пластинки, що жорстко згуртовується з кальцифікованим шаром глибокої зони суглобного хряща. До цього часу показник проліферації хондроцитів у проміжній зоні різко скорочується; продукування компонентів матриксу хондроцитами і їхня репродукція обмежені потребами структурної самопідтримки суглобного хряща. Але гіпертрофовані хондроцити глибокої зони, включаючи і її кальцифікований шар, зберігають життєздатність і продовжують виконувати роль шляхів транспорту живлення від кровоносних капілярів підлягаючої кісткової пластинки до кліток проміжної зони.
Джерела трофіки суглобного хряща варто розглянути докладніше. Розбіжності по цьому непростому питанні породжені і підтримуються в основному двома обставинами: по-перше, відсутністю в суглобному хрящі власних кровоносних судин; по-друге, унікальною топографією хрящового покриву в суглобах, при якій його підстава органічно спаяна з підлягаючою кістою, а вільна поверхня омивається синовіальною рідиною. Саме незаперечний факт прямого контакту суглобного хряща із синовією використовується багатьма як аргумент на користь того, що синовіальна рідина служить джерелом його живлення (Гитис, 1949; Косицин, 1949; Шакалів, 1949; Хем, Кормак, 1983; Павлова й ін., 1988; Дедух і ін., 1992 і ін.). Якщо ж розглядати ситуацію більш об'єктивно, то відразу виникає питання: чи може взагалі харчуватися тихорєцька система з боку поверхні, що постійно піддається деградації і зносу? Тим більше, що на роль джерела трофіки пропонується середовище - синовіальна рідина, що по своєму складі і властивостям ніяк не може бути живильним субстратом. Синовіальна рідина, як відомо, крім діартрозів утримується також у синовіальних сухожильних піхвах. Тут, як і в синовіальних суглобах, вона є ефективним біологічним змащенням, і ніхто не вважав за можливе приписувати їй роль джерела живлення сухожилля. І це зрозуміло, тому що і самому сухожиллі і його оболонці (перитендинеум) усе-таки постачені, хоча і бідно, власними кровоносними судинами. У суглобному хрящі судин ні, як немає судин і в гіаліновому хрящі інших локалізацій, проте живлення останніх однозначне зв'язують із кровоносними судинами прилежащих тканин. Таким джерелом для суглобного хряща є добре васкуляризирована підлягаюча кісткова пластинка, з яким суглобної хрящ структурно об'єднаний у єдине ціле. Правда, цьому єдиному цілому не завжди дається правильна оцінка. Справа в тім, що, як уже згадувалося вище, гіаліновий матрикс у самих нижніх шарах глибокої зони суглобного хряща наповнений кристалами фосфорно-кальцієвих солей, а хондроцити в процесі дозрівання переходять тут у стан набрякання (гіпертрофії). І перше, і друге, по помилковій оцінці деяких дослідників (Хем, Кормак, 1983), неодмінно повинні приводити до загибелі хондроцитів і переходові кальцинованого шару хряща в неживий стан. У такий спосіб на границі суглобного хряща з підлягаючою кістою начебто б створюється перешкода, що виключає можливість дифузії кисню і живильних речовин у хрящ із судин підлягаючої кісти. У дійсності такої перешкоди тут ні, тому що протягом всього індивіда життя суглобної хрящ, у тому числі і клітки в його кальцинованому шарі, зберігають життєздатність завдяки монолітному злиттю з добре васкуляризированою кісткою. Життєздатність хондроцитов у глибокій зоні суглобного хряща підтверджується, зокрема, можливістю їхньої репродукції; тут серед гіпертрофованих хондроцитів зустрічаються клітки з фігурами мітотичного розподілу.
У процесі диференціювання набрякання зрілих хондроцитів сполучене з нагромадженням у їхній цитоплазмі макромолекулярних агрегатів глюкозаміногліканів і глікопротеїдів, що відрізняються високою спорідненістю до полярних молекул води і ковалентно зв'язують них у досить великій кількості. Тому цитоплазма в таких клітках знаходиться в стані гелю, що утримує більш 85% води. Така живаючи гідрофільна "начинка" у клітках ставить них у положення посередників ефективного шляху дифузії в хрящ живильних речовин і кисню.
Поверхнева зона суглобного хряща виключає таку можливість в основному по двох причинах. По-перше, хондроцити тут, як уже згадувалося вище, послідовно (від середньої зони до суглобної поверхні) через недостатню трофіку унаслідок видалення від підлягаючої кісти, деградують і піддаються в кінцевому рахунку повної деструкції. По-друге, із усіх компонентів, що входять до складу синовіальної рідини, у матрикс хряща фізично здатна проникати тільки гіалуронова кислота, що у принципі не може бути субстратом для живлення кліток. До того ж таким шляхом не може надходити до хондроцитів кисень. І все-таки, володіючи властивостями гідрофільності, гіалуронова кислота сприяє підтримці в поверхневій зоні хряща необхідного тургору, створюючи при цьому високий рівень внутрітихорєцького ацидозу. Таким чином, у ланках кістяка, що зчленовуються, з боку порожнини суглоба синовіальним середовищем постійно підтримуються особливі умови, що цілком блокують поширення енхондрального заміщення на суглобної хрящ. Тобто , для кістякових ланок, що зчленовуються, синовіальна рідина в суглобах є не тільки біологічним змащенням тертьових поверхонь, але і необхідною умовою диференціювання на визначеному етапі онтогенезу суглобного хряща і його наступного збереження. З цього, однак, випливає, що якщо з якихось причин така умова порушується, неминуче повинні наступити зміни в стані хрящового покриву суглоба. І дійсно, як було доведено в спеціальних досвідах на собаках (Дружинін, 1889), при вичленовуванні кісти в суглобі з видаленням капсули суглоба із синовіальною оболонкою (тобто джерела продукування синовії), у перші ж дні після операції в глибокому шарі суглобного хряща активізується енхондральна резорбція, і незабаром суглобної хрящ цілком заміщається кісткою.
Якщо звернутися до порівнянь, то можна побачити, що в нижчих хребетних (безхвості амфібії) протягом усього життя а у вищих (ссавці) у ранньому онтогенезі епіфізи довгих кіст і коротких кіст знаходяться в хрящовому стані; тут немає ще структури, що прийнято називати суглобним хрящем. У ссавців у кожнім такому епіфізі й у хрящових закладках коротких кіст ще до початку заміщення кістою утримується своя внутріорганна система кровоносних судин, що харчує всю хрящову масу аж до суглобної поверхні. Система судин, як це видно на проясненому препараті, розміщена усередині хрящових каналів, галузистих по деревовидно-кінцевому принципі. Кожен хрящовий канал несе в собі ажурну мережу кровоносних капілярів у вигляді своєрідної муфти, що зовні нагадує в`язану панчоху. У кінцевих відгалуженнях каналів поблизу суглобної поверхні ажурна судинна муфта закінчується термінальним капілярним ковпачком. Після повного заміщення хрящового епіфіза (закладки короткої кісти) кістою система кінцевих капілярних терминалей майже в незміненому виді зберігається безпосередньо під глибоким шаром суглобного хряща. Тобто, як у хрящовому стані, так і після заміщення кістою всі частини епіфіза (короткої кісти), включаючи і суглобної хрящ, живиться за допомогою єдиної внутріорганної системи крім кровоносних судин. Її термінальні ділянки протягом всього індивіда життя зберігають зв'язок з тією частиною провізорної структури кісткової ланки, що не замістилась кісткою, і перейшла в дефінітивний стан у вигляді суглобного хряща. Можна ще раз повторити ту непорушну істину, що хрящовий покрив в синовіальних суглобах є органічною частиною кісткової ланки і має загальне джерело живлення зі своєю кістковою основою. З боку тертьової поверхні суглобної хрящ постійно несе втрати клітин і міжклітинної речовини. Фізіологічне самовідновлення структури суглобного хряща здійснюється механізмами інтерстиціального росту завдяки проліферації, що підтримується, хондроцитів у проміжній зоні.
Висновок
Хрящова тканина являє собою спеціалізований різновид сполучної тканини, яку відносять до групи тканин з опорною та формоутворюючою функцією. Основні компоненти хрящової тканини — міжклітинний матрикс та клітини — хондроцити. До матриксу входять волокна та основна міжклітинна речовина.
Хрящова тканина безхвостих земноводних характеризується подібністю в будові з вищими хребетними тваринами. У дорослих особин безхвостих земноводних у деяких суглобах кінцівок (наприклад, у колінному) кінці довгих трубчастих кістках протягом всього життя зберігають хрящовий стан. У той же час в інших кістякових ланках (наприклад, у лопатко-плечевому суглобі) хрящові епіфізи в основній своїй масі заміщаються кісткою. Тільки вузький суглобний край залишається хрящовим, за аналогією його можна порівняти із суглобним хрящем ссавців. У першому випадку можна говорити лише про суглобні поверхні на сполучених хрящових епіфізах. В другому випадку маємо справу зі збереженої з боку суглоба і периферичною частиною хрящового епіфіза, що перейшла в дефінітивне, стан, що прийняв форму і функцію самостійного компонента - суглобного хряща.
Список літератури
Александровская О.В., Радостна Т.П., Козлов НА. Цитология, гистология и эмбриология. - М.: ВО «Агропромиздат», 1987 - 448 с.
Белоусов Л. В. Введение в общую эмбриологию: Изд-во Московского университета, 1980. — 210 с.
Бодемер Ч. Современная эмбриология. — М.: «Мир», 1971. — 446 с.
Вельш У., Шторх Ф. Введение в цитологию и гистологию животных. — М.: «Мир», 1972. — 259 с.
Иванов И.Ф., Ковальский П.А. Цитология, гистология, эмбриология. М.: «Колос», 1976. — 447 с.
Международная гистологическая номенклатура / Под ред. Ю.И.Копаева — К., 1980. - 113 с.
Свенсон К., Уэбстер П. Клетка. — М.: «Мир», 1980. — 303 с.
Серов В.В., Шехтер А.Б. Соединительная ткань. — М.: «Медицина», 1981. — 312 с.
Токин Б.П. Общая эмбриология. М.: «Высшая школа», 1977. — 509 с.
Хэм А., Кормак Д. Гистология. — Т. 1—5. — М.: «Мир», 1983. — 272 с.
Хрящ / Павлова В.М., Копьева Т.Н., Слуцкий Л.И., Павлов Г.Г. — М.: «Медицина», 1988. — 317 с.
Ченцов Ю.С. Общая цитология: Изд-во МГУ, 1984. — 350 с.
Шмидт Г.А. Эмбриология животных. Ч.1: Общая эмбриология: Государственное изд-во «Советская наука». — М., 1951. — 354 с.

Нравится материал? Поддержи автора!
Ещё документы из категории биология:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ