Статья «Дистанция вспугивания сизых голубей»
«Дистанция вспугивания сизых голубей».
СОДЕРЖАНИЕ
ВВЕДЕНИЕ
1.1. Постановка проблемы (из статьи)
Цели и основные задачи исследования
МАТЕРИАЛ и МЕТОДИКА
2.1. Места и сроки проведения исследований
2.2. Объём собранного материала
2.3. Методика исследований (из статьи)
2.4 Теоретическая часть
ЗАКЛЮЧЕНИЕ и ВЫВОДЫ
Приложение: таблицы
Результаты и выводы
ЛИТЕРАТУРА
Введение.
Оценка толерантности птиц к человеку и движущемуся автотранспорту
Город, как среда обитания диких животных отличается от природной среды рядом особенностей. В первую очередь разнообразием местообитаний и их мозаичностью (Клауснитцер, 1990), постоянным возрастанием разнообразия предметных компонентов, наконец, присутствием человека, как весьма значимого фактора (раздражителя), влияющего на поведение животных, обитающих в городе (Мешкова, Федорович, 1996). Городская среда (в особенности селитебный ландшафт) характеризуется высоким уровнем антропогенного беспокойства. В городской среде преимущество имеют те популяции птиц-синурбанистов, которым присуща большая степень антропотолерантности.
Критерием при оценке степени непосредственной толерантности птиц к фактору беспокойства традиционно служит, «дистанция вспугивания» (ДВ) (Владышевский, 1975; Вахрушев, Зюзин, 1984; Резанов А.А., 2002, 2003, 2006; Резанов, Резанов, 2002). Иногда её определяют как «дистанцию взлёта» (“flight-initiation distance”) – т.е. дистанцию, с которой птица взлетает при приближении хищника (Dill, Houtmab, 1989; Kramer, Bonenfant, 1997; Metcalf et al. 2000). Мы считаем термин ДВ более удачным, поскольку он подразумевает более широкий спектр локомоций и форм защитных реакций птицы при приближении опасности, а не только взлёт.
В качестве модельного вида для выявления реакции птиц на пешеходов и автотранспорт, нами выбрана серая ворона (Corvus cornix), как вид доступный для наблюдений в течение всего года и встречающийся в местообитаниях с различным уровнем фактора беспокойства. Дистанция вспугивания серой вороны оценивалась нами при её нахождении в «наземном измерении» (Резанов, 2006) в зоне прямых контактов с человеком и автотранспортом.
Оценка реакции серой вороны на человека
В поведении серой вороны при появлении сигнала опасности органично сочетается как реакция затаивания (реакция затаивания представляется наиболее консервативной), так и реакция вспугивания, что также подтверждает их высокую поведенческую пластичность. Д.В. Владышевский (1975) подобные реакции характеризовал, как видоспецифические генерализованные защитные реакции, которые у синантропных видов, претерпевают существенную дифференцировку, выражающуюся в правильной оценке поведения человека в каждой конкретной ситуации.
ДВ птиц обусловлена многими факторами: видоспецифическими и индивидуальными особенностями птицы, характером и направленностью движения человека, возрастной категорией людей, многолюдностью и характером биотопа, соотношением частоты отрицательных и индифферентных контактов с человеком (Крушинский, 1958; Владышевский, 1975; Константинов, Бабенко, Барышева, 1982; Вахрушев, Зюзин, 1984; Резанов А.А., 2002; Резанов, Резанов, 2002).
Л.В. Крушинский (1958), на примере оборонительной реакции (термин ДВ автор не использовал) серой вороны описал явление экстраполяции, заключающееся в том, что «животные могут реагировать не только на местоположение находящегося в данный момент раздражителя, но и на то место, в котором этот раздражитель будет находиться через определённое время при его закономерном движении». Было обнаружено, что если движение человека направлено мимо вороны, то птица подпускает его на заметно меньшее расстояние (36 шагов – наблюдения проведены на открытой местности – А.Р.), нежели при его движении прямо на птицу (49 шагов). Следовательно, для проявления защитной реакции имеет значение не столько близость раздражителя, сколько направление его движения.
Показатель ДВ носит строго индивидуальный характер и формируется как усреднённая сбалансированная реакция на различную степень опасности со стороны объекта представляющего потенциальную или реальную угрозу, каким и является человек. И выигрывают те птицы, у которых этот показатель отличается наибольшей адекватностью в каждом конкретном случае. C.J. Protor et al. (2001), используя математическое моделирование показал, что птицы, взлетающие при любом предупреждающем сигнале о потенциальной опасности, с энергетической точки зрения существенно проигрывают тем особям, которые не реагируют вхолостую, а взлетают только при реальной опасности, т. е. подтверждённой множественными предупреждающими сигналами.
Началом адекватной защитной (оборонительной) реакции птиц на фактор беспокойства со стороны человека можно считать любую смену рода активности, в т.ч. на уровне элементарных двигательных актов.
Весь спектр защитных реакций вороны на приближающегося человека в зависимости от степени настороженности, мы ранжировали на 5 показателей, выраженных в баллах:
видимая реакция отсутствует (0);
Реакция настороженности: сканирование (поворот головы в сторону наблюдателя) и т.п. (0,5);
приседание (1);
отход или отпрыгивание в сторону от наблюдателя (2);
взлёт (3).
Дистанция вспугивания определялась при демонстрации человеком индифферентного отношения к птице во время его подхода к ней. При регистрации дистанции вспугивания мы оценивали направление движения человека относительно птицы: 1) прямой подход: оценка дистанции вспугивания при движении объекта прямо на птицу (вспугивание на момент нахождения птицы прямо от объекта); 2) непрямой подход: а) оценка дистанции вспугивания по перпендикуляру (вспугивание в момент нахождения птицы сбоку от объекта, когда объект поравнялся с ней; дистанцией вспугивания в этом случае является перпендикуляр от трансекты движения объекта к точке нахождения птицы); б) оценка дистанции вспугивания по гипотенузе (вспугивание птицы до того момента, как объект предположительно поравняется с ней; дистанция вспугивания в этом случае является гипотенузой условного треугольника, один катет которого – расстояние от объекта до точки начала перпендикуляра, а другой катет – сам перпендикуляр) (Рис. 1-2).
Показательно, что при отсутствии прямого преследования со стороны человека и достаточно индифферентном его отношении к птицам, ДВ и интенсивность прохождения людей находятся в прямой зависимости друг от друга.
На основе существенных различий в уровне фактора беспокойства со стороны пешеходов, мы условно выделили 3 типа местообитаний: 1) жилые микрорайоны - ЖМ; 2) дорожки в парках - ДП; 3) открытые пространства (ОП) без дорог или с неразвитой сетью дорог - луга, овраги, пустыри. Фактор беспокойства (число пешеходов и т.п.) учитывался в различных местообитаниях. В целом, взято 290 проб по 15 мин каждая. Получены следующие показатели: ОП – 6.12 чел./15 мин (n = 77); ДП – 8.24 (n = 82); ЖМ – 12.5 (n = 131). Исходя из соотношения 6.12:8.24:12.5, введены коэффициенты 1:1.35:2.04, использованные при статистической обработке материала.
Оценка фактора беспокойства проводилась методом учёта интенсивности движения людей (пешеходы, велосипедисты и пр.) в местах, где определялась ДВ. Подсчеты проводились периодически в течение 15-20 мин в различные дни недели (будни, выходные) и в различное время суток (утро, день, вечер) – в соответствии со сроками оценки ДВ. Для выбранных типов местообитаний рассчитаны средние показатели фактора беспокойства. Максимальные показатели фактора беспокойства (принято, что именно максимальный показатель фактора реально определяет ДВ птицы в данном местообитании) переведены в условные единицы – баллы, использованные при статистической обработке собранного материала.
По-видимому, средняя величина ДВ может также служить объективным критерием оценки уровня фактора беспокойства в конкретной местности.
По исследованиям в различных районах Москвы получена однозначная генеральная тенденция, показывающая, что с ростом фактора беспокойства (от 1 до 2.04) ДВ серой вороны снижается (r = - 0.5284; P < 0.001; n = 1053). Чёткое соответствие между ДВ и величиной фактора беспокойства видно из следующих расчетов. Средние ДВ по местообитаниям составили: для ЖМ – 2.51 м (n = 323); для ДП – 4.50 м (n = 315); для ОП – 9.17 м (n = 415). Соответствующие показатели относятся друг к другу как 1 : 1.79 : 3.65. Таким образом, сравнение показателей по ДВ и фактору беспокойства, со всей очевидностью, демонстрируют обратную зависимость (Табл. 1).
Таблица 1
Общая оценка зависимости между дистанцией вспугивания серой вороны и величиной фактора беспокойства
Оцениваемые показатели
Расчетные коэффициенты
Жилые микрорайоны (ЖМ)
Дорожки в парках (ДП)
Открытые пространства (ОП)
Фактор беспокойства
2.04
1.35
2.04
Дистанция вспугивания
1
1.79
3.65
Исходя из известных различий в составе населения ворон Москвы в зимний и летний периоды (Ильичев, Бутьев, Константинов, 1987), мы провели в отдельных «точках» города сравнение ДВ по сезонам. Были получены соответствующие данные для ЖМ и ОП. В зимний период в городе сосредоточено весьма разнородное население вороны: собственно «городские» (оседлая популяция) и «пришлые», прилетевшие на зиму в город из его окрестностей и обширных территорий, находящихся к северо-востоку от столицы. Соответственно, мы предположили, что зимой ДВ серой вороны и отклонения от средней арифметической (± S.E.) будут выше, чем летом. Предположение, на уровне тенденции (td = 0.25 и td = 0.15, для ЖМ и ОП, соответственно), подтвердилось. По ЖМ ДВ вороны зимой составила 3.79 ± 2.71 м (lim 0.4 -12.5; n = 39; P = 0.001), а летом - 3.07 ± 1.02 м (lim 0.8-8.0; n = 60; P = 0.001); по ОП - 12.81 ± 5.29 м (lim 2.5-31; n = 35; P = 0.001) зимой и 11.86 ± 3.10 м (lim 5-20; n = 24; P = 0.001) летом.
Выявленная закономерность подтверждает положение, высказанное другими авторами ранее (Вахрушев, Зюзин, 1984), согласно которому с ноября в Москве изменчивость ДВ серой вороны должна возрастать, ввиду притока птиц их других районов. Мы также условно оценили разнообразие населения серой вороны зимой (2 балла) и летом (1 балл) и сравнили посезонное изменение ДВ в ЖМ и ОП. Полученные тенденции совпадают с предыдущими расчетами: зимой ДВ и отклонения от средней арифметической (± S.D.) у вороны были выше. И, в то же время, выявленные различия также статистически недостоверны: y = 0.7104x + 2.3613 (R2 = 0.0177, P>0.05, n = 99) и y = 0.6482x + 11.189 (R2 = 0.0028, P>0.05, n = 59) для ЖМ и ОП, соответственно.
В ЖМ, даже зимой сосредоточены только «городские» вороны, не совершающие суточных кормовых перелетов и остающихся ночевать в микрорайоне. На ОП - в основном, «пришлые»; в частности, в Коломенском, где проведены основные наблюдения, вороны, за исключением одиночек, не ночуют. Исходя из этого, мы ожидали, что разброс показателей ДВ у ворон из ЖМ и ОП зимой должен быть выше, чем летом. Действительно, зимой достоверность различий ДВ у ворон исследуемых территорий оказалась ниже (td = 1.52), чем летом (td = 2,70). Также, в ноябре-марте показатель ± S.E. в пределах ЖМ и ОП отличался большей амплитудой, чем в мае. Аналогичная картина получена при сопоставлении уравнений регрессии для зимы (y = - 0.9671x + 13.453; R2 = 0.3958, P<0.001, n = 74) и для лета (y = - 0.974x + 12.811; R2 = 0.6629, P<0.001, n = 84).
Однако, различие средних показателей ДВ ворон в зимний и летний периоды, вероятно вызвано не столько притоком в город зимой «пришлых» ворон, а сколько общем снижением интенсивности прохождения людей в этот период, что соответственно приводит в среднем к увеличению ДВ. Данное предположение основано на том, что ввиду высокой пластичности поведения вида в целом, «пришлые» вороны привыкшие к определённой встречаемости людей в летнем местообитании, тем не менее достаточно быстро адаптируются к уровню фактора беспокойства, соответствующему их местообитанию на зимней период времени.
Можно предположить, что величина ДВ у ворон, как птиц с высоким уровнем пластичности поведения, скорее определяется конкретной ситуацией, чем их принадлежностью к определённым внутрипопуляционным группировкам. Каждая птица ведет себя адекватно условиям, в которых она оказывается. Так, вороны ОП довольно чутко реагируют на приближение человека, не подпуская его на близкое расстояние, и в то же время, почти вплотную подходят к людям во время пикников в надежде поживиться чем-нибудь съестным. Более того, как показывают наблюдения в Коломенском и Нескучном, сравнительно пугливые «парковые» вороны, абсолютно доверчивы к людям, которые их подкармливают.
Нами проведена оценка зависимости между дистанцией по трансекте движения, по гипотенузе (реальная ДВ) и перпендикуляром (т.е. расстоянием по прямой от трансекты). В этих ситуациях отмечена сходная тенденция: рост ДВ при снижении расстояния по перпендикуляру (r = -0.2195 и r = - 0.21; P < 0.001; n = 1053). Иными словами, более прямой подход приводит к взлёту птиц с большего расстояния.
Показано, что с ростом числа птиц в группе, ДВ растёт (r = 0.15; P < 0.001; n = 1053). Возможно, это происходит по причине стимуляции сравнительно спокойных особей более беспокойными птицами. Хотя неоднократно приходилось наблюдать разновременный взлёт ворон даже в больших группах. При этом птицы демонстрировали самые различные ДВ.
Мы также попытались оценить зависимость степени реакции (от 0 до 3 баллов) от дистанции между птицей и наблюдателем. Степень реакции птиц на внешний раздражитель при сокращении расстояния с этим раздражителем, естественно усиливается: от моторных актов (сканирование головой, приседание) до актов локомоторных (отскок, отход, взлёт). Полученная нами тенденция (r = 0.2768; P < 0.001; n = 1053) свидетельствует об обратном: увеличение дистанции между птицей и человеком ведёт к росту уровня реагирования со стороны птицы. Данный нонсенс, по-видимому, можно объяснить следующим образом: 1) за счёт значительного числа наблюдений на ОП (это видно из графика), где птицы отличаются пугливостью и взлетают с больших расстояний; 2) в большинстве случаев птицы находились на прямой движения человека (т.е. осуществлялся прямой подход) и взлёт был их естественной реакцией. В то же время обнаружена довольно чёткая статистически достоверная тенденция (r = - 0.2953; P < 0.001), согласно которой с ростом расстояния по перпендикуляру степень проявления реакции у ворон снижается вплоть до нуля, когда птица на человека не реагирует. Отмеченная тенденция видна также из средних показателей величины перпендикуляра (Табл.2).
Таблица 2
Зависимость степени проявления реакции птицы
от характера подхода к ней наблюдателя
Степень реакции птицы, баллы
Число подходов
Показатель (Х) величины перпендикуляра
0
106
3.34
1
42
2.66
2
184
1.21
3
721
1.10
Известно, что в безлюдных местах (особенно там, где нет охоты на птиц) некоторые виды совсем не пугливы. Увеличение частоты «настойчивых» подходов к птицам способствует увеличению ДВ (Владышевский, 1975), в то время как на многолюдных улицах она сокращается (Вахрушев, Зюзин, 1984). Вероятно, в последовательности «безлюдные районы…- многолюдные улицы» прослеживается закономерность: с ростом фактора беспокойства, особенно, если он сопровождается прямым преследованием, ДВ растет до определенного предела и стабилизируется. В населенных пунктах, особенно в городах, с ростом фактора беспокойства (не подкрепляемого преследованием) ДВ начинает неуклонно падать, доходя до допустимого минимума у урбанизированных популяций птиц.
Показано, что в других городах и населённых пунктах ДВ серой вороны заметно выше (Владышевский, 1975; Родимцев и др., 1984), чем у московских птиц (Константинов, Бабенко, Барышева, 1982; Вахрушев, Зюзин, 1984; Резанов, Резанов, 2002). В частности, в Новокузнецке ДВ серой вороны составила 15-20 м (Родимцев и др., 1984), а на Украине – 12.4 м на дорогах и 34.6 м – в стороне от дорог (Владышевский, 1975). Можно предположить, что, в целом, ДВ зависит от целого ряда показателей: от степени урбанизации популяции, от особенностей городской среды обитания, от характера и уровня фактора беспокойства. Без знания конкретных цифр, характеризующих отмеченные показатели, дать корректное объяснение выявленных заметных расхождений в ДВ различных популяций серой вороны не представляется возможным.
Важно отметить то, что одна и та же ворона, находясь в различных биотопах, изначально проявляет ту степень толерантности (выражается в разбросе показателей индивидуальной ДВ), которая выработалась у неё в данном местообитании, исходя из предшествующего опыта. Данная степень толерантности является результатом совокупного действия всех факторов обуславливающих ДВ. Соответсвующую каждому конкретному местообитанию степень толерантности, можно сравнить с ответной реакцией на сложный стимул (сигнальный комплекс), которым и является этот биотоп с сопутствующими ему факторами, влияющими на толерантность. В сознании птицы возникает связь между биотопом и этими факторами.
Однако сама степень толерантности формируется на базе конкретного биотопа в результате процессов привыкания к человеку. Степень толерантности может меняться в случае изменения интенсивности прохождения людей или соотношения частот отрицательных, индифферентных и положительных контактов с человеком, а также по причине появления новых факторов сопутствующих человеку (например, выгул собак) и изменения характера самого ландшафта.
Процесс привыкания может прекратиться при снижении интенсивности прохождения людей и увеличении частоты отрицательных контактов с людьми, а также при появлении новых факторов сопутствующих человеку, т.е. исходная оборонительная реакция на человека, как раздражитель, будет восстанавливаться и ДВ соответственно увеличится. К незначительному временному увеличению ДВ также может приводить изменение характера ландшафта, т.к. с появлением новых элементов снижается идентификация местности, она становится малознакомой и у птиц возрастает доля оборонительных реакций на многие сопутствующие данной местности показатели (неофобия), в том числе и на человека.
Наоборот, к общему уменьшению ДВ ведёт усиление процесса привыкания к человеку (увеличение интенсивности прохождения и встречаемости людей, увеличение частоты индифферентных контактов с человеком) и оценка человека, как положительный стимул (увеличение частоты положительных контактов с человеком, например, в результате подкормки).
Процесс привыкания может, как прекращаться (при изменении показателей или появлении новых факторов), так и восстанавливаться (при возвращении прежних показателей), что обуславливает возможность изменения ДВ у птиц.
Индивидуальная ДВ у каждой птицы постоянно варьирует в зависимости от ситуации. По нашим наблюдениям, при приближении человека к группе из нескольких и более ворон, реакция вспугивания последних чаще всего асинхронна, в отличие от уток (Anas spp.), которые, как правило, вовлекаются в реакцию бегства в радиусе 10-25 м (Авилова и др., 1994). Это различие объясняется тем, что в поведении ворон ярче выражены их индивидуальные свойства. Птица реагирует прежде всего на действия человека, оценивая их, а не на реакцию своих сородичей. При приближении человека, дистанция от него до каждой птицы в группе будет различной и, по-видимому, для птиц находящихся от него дальше будет нецелесообразно сразу покидать место, следуя примеру других особей, т. к. он может кардинально изменить направление своего движения. Исходя из вышесказанного, можно заключить, что реакция серой вороны на фактор беспокойства со стороны человека отличается наибольшей адекватностью, вследствие чёткой дифференциации её защитных реакций.
В столь высокой адаптивности поведения вороны играет значительную роль присущий им высокий уровень развития элементарной рассудочной деятельности (Зорина, 1984 и др.).
Успешность внедрения врановых в определённые биотопы антропогенного, в частности селитебного ландшафта и их успешная поведенческая адаптация к условиям жизни в них, во многом обусловлена, как быстрым привыканием к создавшимся относительно стабильным условиям окружающей среды в конкретных биотопах, так и эффективной адаптацией к постоянным незначительным (фоновым) изменениям этой среды.
Адаптация к резким значительным и непредсказуемым изменениям, присущим городской среде обитания, проходит тяжелее и носит более длительный характер, в связи с тем, что предугадать их невозможно. Тем не менее, у птиц, входящих в состав городских популяций, в полной мере освоивших город, как среду обитания, по всей видимости, выработалась некая предрасположенность к адаптации в условиях подобных значительных и непредсказуемых изменений и определённая степень готовности к ним.
Реакция птиц на движущийся автотранспорт
Глобальное расширение сети автодорог позволило некоторым зоологам рассматривать их как самостоятельную экологическую систему. Вопросу о роли дорог в жизни птиц посвящено немало работ, среди которых много внимания уделено проблеме гибели птиц на автодорогах (Нанкинов, Тодоров, 1983; Телегин, Ивлева, 1983; Рахилин, 1996; Лысенков и др., 2000), а также питанию птицами, сбитыми животными (Guillory, Le Blane, 1975; Лысенков и др., 2000) и особенностям кормового поведения птиц, связанного с движущимся автотранспортом (Резанов, 1998 и др.).
Разыскивая корм на автодорогах, птицы сталкиваются с ситуацией, у которой нет биологических аналогов – с ситуацией, когда к птице приближается потенциальный «хищник» (объект, в котором заключена опасность для жизни – в данном случае автомобиль) на скоростях, нередко превышающих биологические возможности реального хищника.
Задачей нашего исследования было оценить степень толерантности серой вороны на приближающийся автотранспорт по дистанции вспугивания. Наблюдения проведены в Москве летом 2001 года. При оценке дистанции вспугивания вороны (n = 113) на приближающийся автотранспорт, наблюдатель, находящийся у дороги, фиксировал следующие основные моменты: 1) тип автотранспорта (легковой - 1, малолитражный - 2, грузовой – 3 балла); 2) скорость автомобиля – по визуальной оценке, по возможности, с точностью до 10 км/ч; 3) величина перпендикуляра от трансекты движения автомобиля к точке нахождения птицы; 4) количество птиц в группе. Попытки проводить наблюдения с движущегося автотранспорта оказались менее удачными. Основное затруднение вызвала оценка дистанции вспугивания, проводившаяся наблюдателем непосредственно из автомобиля.
Для сравнения также оценены ДВ сизого голубя, галки, домового воробья. В предлагаемой таблице (Табл. 3) приведены данные по ДВ различных видов птиц при приближении автотранспорта. Обращает на себя внимание, что величина ДВ у врановых птиц заметно выше, чем у сизого голубя и домового воробья. Возможно, врановые птицы более адекватно оценивают ситуацию на дорогах, ту опасность, которую представляет собой движущийся (приближающийся) автотранспорт.
Таблица 3
Дистанция вспугивания птиц при приближении автотранспорта (г. Москва)
Виды птиц
Количество вспугиваний
_
Среднее значение (X) дистанции вспугивания, м
Lim
Сизый голубь
129
4.28
0.5-11
Серая ворона
113
9.72
0.5-60
Галка
19
13.36
4-27
Домовой воробей
63
4.91
2-10
Проведён сравнительный анализ оценки дистанции вспугивания у птиц в зависимости от типа и скорости приближающегося автотранспорта, в зависимости от величины перпендикуляра от точки нахождения птицы к трансекте движущегося автомобиля, в зависимости от числа птиц в группе (Табл. 4).
1) Зависимость дистанции вспугивания птиц от типа автотранспорта. Сам по себе тип автотранспорта не таит в себе никакой опасности для птиц, но его движение в сочетании с размерами и шумовыми эффектами, по-видимому, могут оказывать на птиц определенное воздействие. Опять же, врановые, на наш взгляд, в данной ситуации, продемонстрировали наиболее адекватную реакцию, не показав никакой зависимости ДВ от типа приближающегося автотранспорта. В то же время, сизый голубь четко реагировал на размеры транспортных средств, особенно на автобусы, увеличением ДВ (r = 0.3291; n = 129; P < 0.001).
2) Зависимость дистанции вспугивания птиц от скорости автотранспорта. Наиболее чёткая зависимость ДВ от скорости приближающегося автотранспорта показана врановыми; серой вороной показана достоверная положительная тенденция (r = 0.5042; n = 113; P < 0.001), а галка показала обратную, но статистически недостоверную зависимость (r = - 0.3489; n = 19; P > 0.05).
3) Зависимость дистанции вспугивания от величины перпендикуляра от точки нахождения птицы к трансекте движения автомобиля. Сизый голубь показал труднообъяснимую зависимость, согласно которой с ростом перпендикуляра дистанция вспугивания, против всякого ожидания, увеличивается (r = 0.6948; n = 129; P < 0.001). Аналогично у домового воробья, но отмеченная тенденция статистически недостоверна (r = 0.2223; n = 63; P > 0.05). Серая ворона показала ожидаемое снижение величины ДВ при росте перпендикуляра (r = 0.2341; n = 113; P < 0.05). Галка не показала никакой тенденции.
4) Зависимость дистанции вспугивания от числа птиц в группе. Наиболее чёткая зависимость показана галкой: рост ДВ с увеличением числа птиц в группе (r = 0.4496; n = 19; P = 0.05); данная тенденция практически на грани статистической недостоверности. Такая же тенденция, но статистически достоверная (r = 0.261; n = 129; P < 0.01), продемонстрирована сизым голубем. По-видимому, наиболее осторожные особи провоцируют взлёт стаи в целом. Домовый воробей и серая ворона не показали видимых тенденций.
Таблица 4
Зависимость дистанции вспугивания у птиц на автодорогах
от различных факторов. Москва
Виды птиц
Зависимость дистанции вспугивания от
Типа автотранспорта
Скорости автотранспорта
Величины перпендикуляра к трансекте движения автотранспорта
Числа птиц в группе
Сизый голубь
Прямая
(r = 0.3291;
P < 0.001)
Прямая
(r = 0.0721;
P > 0.05)
Прямая
(r =0.6948;
P < 0.001)
Прямая
(r = 0.261;
P < 0.01)
Серая ворона
Не выражена
Прямая
(r = 0.5042;
P < 0.001)
Обратная
(r = 0.2341;
P < 0.05)
Не выражена
Галка
Не выражена
Обратная
(r = - 0.3489; P > 0.05)
Не выражена
Прямая
(r = 0.4496;
P = 0.05)
Домовый воробей
Прямая, незначительная
(r = 0.1049;
P > 0.05)
Не выражена
Прямая
(r = 0.2223;
P > 0.05)
Не выражена
Целью нашего исследования является выявление действия антропогенного фактора на сизых голубей.
Материал и методика.
Места и сроки проведения исследований.
Наблюдения проводились в Москве и Московской области, на протяжении года, что бы выявить влияние внешних факторов на ДС сизых голубей.
Объем собранного материала.
В ходе исследований было собрано 80 наблюдений за разными сизыми голубями: одиночными особями, особями в группах, особями манипулирующими и так же ни чем не занимающимися в начале исследования.
Методика исследований.
Городская среда (в особенности селитебный ландшафт, рассматриваемый как ключевой признак города) характеризуется высоким уровнем антропогенного беспокойства. В городской среде преимущество имеют те популяции птиц-синурбанистов, которым присуща большая степень антропотолерантности.
Условно можно выделить два уровня антропотолерантности:
Общая, или фоновая - она в той или иной степени присуща всем птицам, приуроченным непосредственно к селитебному ландшафту, адаптированным к фоновому воздействию города.
Частная, или непосредственная (контактная)- присуща птицам, вступающим в прямые контакты с человеком, а также с мобильными наземными элементами городской среды, имеющими антропогенное происхождение: движущийся транспорт, работающая техника и др.
Можно также выделить 2 вида измерений, показывающих существование птиц относительно человека и мобильных наземных элементов антропогенного происхождения:
Наземное измерение. В нем существуют птицы, которые значительную часть времени находятся на «земле» (наземный субстрат или водная поверхность) в «измерении человека» и мобильных наземных элементов антропогенного происхождения и вступают с ними в прямые контакты. Это типичные синантропы: сизый голубь, серая ворона, галка, грач, кряква, воробьи и др.
Надземное измерение. В нем существуют птицы, которые в силу своих биологических особенностей значительную часть времени находятся над землей в ином измерении, относительно человека и мобильных наземных элементов антропогенного происхождения и не имеют с ними прямых контактов. Например, черный стриж, ласточки, обыкновенная пустельга и др.
. Расхождение птиц по измерениям зависит от среды, находясь в которой они разыскивают корм: птицы «надземного измерения» предпочитают разыскивать пищу находясь в воздушной среде, а птицы «наземного измерения» - на твердом субстрате. Контакт с человеком и иными наземными элементами, птиц обоих измерений, возможен также в период их гнездования или отдыха в надземном измерении (орографические сооружения, деревья и т.п.), где и проходит условная граница измерений. При этом многие птицы «надземного измерения», гнездясь или отдыхая на определенном высотном уровне, избегают подобных контактов.
Отсюда очевидна неправомерность сравнения антропотолерантности чёрного стрижа и сизого голубя, находящихся относительно человека в разных «измерениях». Фоновой толерантностью обладают соответственно птицы обоих измерений, а непосредственная толерантность проявляется у птиц преимущественно «наземного измерения». Фоновая толерантность служит качественным критерием антропотолерантности, а непосредственная толерантность - количественным показателем.
Критерием при оценке степени непосредственной толерантности птиц к фактору беспокойства традиционно служит «дистанция вспугивания» (Владышевский, 1975; Вахрушев, Зюзин, 1984; Авилова и др. 1994; Резанов А.А., 2002, 2003). Иногда её определяют как «дистанцию взлёта» (“flight-initiation distance”) (Dill, Houtmab, 1989; Kramer, Bonenfant, 1997; Metcalf et al. 2000). Мы считаем термин дистанция вспугивания более удачным, поскольку он подразумевает более широкий спектр локомоций при приближении опасности, а не только взлёт. Вместе с тем, дистанция вспугивания подразумевает достаточно явные и чёткие формы защитных реакций.
Нами предложена оригинальная методика оценки антропотолерантности птиц. Весь спектр защитных реакций птиц на приближающегося человека и автотранспорт, ранжировали по четырём различающимся показателям: 1) отсутствие видимой реакции; 2) приседание; 3) отскок или отход; 4) взлёт. «Дистанция вспугивания» определена как дистанция проявления у птицы локомоторной реакции (наземной или воздушной локомоции), направленной на уход от раздражителя, при приближении к птице человека, автотранспорта и т.д. Дистанция вспугивания определялась при демонстрации человеком индифферентного отношения к птице во время его подхода к ней. При регистрации дистанции вспугивания мы оценивали направление движения человека или автотранспорта относительно птицы: 1) прямой подход: оценка дистанции вспугивания при движении объекта прямо на птицу (вспугивание на момент нахождения птицы прямо от объекта); 2) непрямой подход: а) оценка дистанции вспугивания по перпендикуляру (вспугивание в момент нахождения птицы сбоку от объекта, когда объект поравнялся с ней; дистанцией вспугивания в этом случае является перпендикуляр от трансекты движения объекта к точке нахождения птицы); б) оценка дистанции вспугивания по гипотенузе (вспугивание птицы до того момента, как объект предположительно поравняется с ней; дистанция вспугивания в этом случае является гипотенузой условного треугольника, один катет которого – расстояние от объекта до точки начала перпендикуляра, а другой катет – сам перпендикуляр).
Показатель дистанции вспугивания птиц обусловлен усреднённым показателем фактора беспокойства. Оценка фактора беспокойства проводилась методом учёта интенсивности прохождения людей (пешеходы, велосипедисты и т.п.) и автотранспорта в различные дни недели (будни, выходные) и время суток (утро, день, вечер) в местах, где определялась дистанция вспугивания. При отсутствии прямого преследования со стороны человека и достаточно индифферентном его отношении к птицам, дистанция вспугивания птиц и интенсивность прохождения людей находятся в прямой зависимости друг от друга. На основе существенных различий в интенсивности фактора беспокойства со стороны пешеходов, мы условно выделили 3 типа местообитаний и рассчитали средние коэффициенты фактора беспокойства: 1) жилые микрорайоны (коэффициент беспокойства 2.04); 2) дорожки в парках (1.35); 3) открытые пространства без дорог или с неразвитой сетью дорог (1.0 – за единицу принята самая низкая интенсивность фактора беспокойства). Баллы отражают максимальные показатели фактора беспокойства в каждом типе местообитания (принято, что именно максимальный показатель фактора реально определяет дистанцию вспугивания птицы в данном местообитании).
В рамках темы проведены наблюдения по оценке дистанции вспугивания целого ряда синантропных птиц пешеходами (Резанов, Резанов, 2002) и движущимся автотранспортом (Резанов А.А, 2005).
Показана однозначная генеральная тенденция: с ростом фактора беспокойства дистанция вспугивания серой вороны снижается (r = - 0.53; P < 0.001; n = 1053). При снижении расстояния по перпендикуляру отмечен рост дистанции вспугивания, т.е. чем подход более прямой, тем с большего расстояния птица реагирует взлётом. Отмечено, что с увеличением расстояния по перпендикуляру степень проявления беспокойства у птицы снижается и полностью исчезает. Зимой дистанция вспугивания серой вороны демонстрирует более высокие отклонения от средней арифметической (но только на уровне статистически недостоверной тенденции), что согласуется с положением, высказанным А.А. Вахрушевым (1984), что с ноября, ввиду притока в город на зимовку птиц из других районов, дистанция вспугивания ворон возрастает.
Показана зависимость дистанции вспугивания ворон от числа особей в группе. Детальный анализ наблюдений за поведением ворон показывает высокую индивидуальность реакции птиц с сохранением общей тенденции к росту дистанции вспугивания (характерно для исследованных видов), свидетельствующей о стимуляции взлёта со стороны наиболее бдительных особей.
Рассчитана дистанция вспугивания 4 видов птиц при приближении автотранспорта. Проведен анализ зависимости дистанции вспугивания от типа и скорости приближающегося автотранспорта, от величины перпендикуляра и от числа птиц в группе. Адекватная реакция отмечена у серой вороны: в частности, птицы не реагировали изменением дистанции вспугивания на разные типы автотранспорта, но чётко увеличивали её при повышении скорости автомобиля. Голуби и галки показали рост дистанции вспугивания от числа птиц в группе, по-видимому, за счёт наиболее осторожных птиц.
Теоретическая часть.
Сизый голубь.

Домен:
Царство:
Тип:
Класс:
Отряд:
Семейство:
Род:
Вид:
Сизый голубь
Си́зый го́лубь (лат. Columba livia) — широко распространённая птица семейства голубиных, родиной которой считаются Европа, Юго-Западная Азия и Северная Африка. Ещё в глубокой древности эти птицы были приручены человеком, в результате были выведены так называемые домашние голуби (Columba livia var. domestica). Во времена Великих открытий человек перевозил с собой всё своё имущество, в том числе и голубей. Впоследствии одичавшие голуби (Columba livia var. urbana) широко распространились в мире и стали синантро́пами, привычными обитателями больших городов и сельскохозяйственных ферм.[1][2]
В дикой природе сизый голубь обычно селится на прибрежных скалах, в горных ущельях либо вдоль обрывистых берегов рек, часто поблизости от зарослей кустарника или сельскохозяйственных угодий. Их полудомашние потомки легко адаптировались к жизни возле человеческого жилья, поскольку каменные постройки напоминают природные места обитания, а пищевые отходы служат надёжной кормовой базой в любое время года. Несмотря на раннее одомашнивание и различные условия обитания, дикие сизые голуби и их городские сородичи свободно скрещиваются друг с другом, что, по мнению орнитологов, может привести к деградации и постепенному вымиранию первых. В природе птицы обычно живут не более трёх-пяти лет, при разведении в домашних условиях часто доживают до 15-летнего возраста, а отдельные особи и до 35 лет. Народные названия птицы — сизак, сизарь. В разговорной речи часто употребляются слова «гуля», «гулька» (отсюда поговорка «с гулькин нос», то есть очень мало).
Внешний вид
Крупный голубь — длина тела 29—36 см, размах крыльев 50—67 см, вес 265—380 г. Оперение густое и плотное, но при этом перья слабо закреплены в коже. Окрас изменчивый, особенно у городских полудиких птиц — всего насчитывают 28 разновидностей окраски, называемых «морфами». Как правило, голова, шея и грудь пепельно-сизые с зеленоватым, желтоватым либо пурпурным металлическим отливом на шее и груди. Такой же отлив может быть выражен на кроющих крыла. Радужная оболочка красная, оранжевая либо золотисто-жёлтая, при этом внутреннее кольцо более бледное. Вокруг глаз имеются участки неоперённой, голубовато-серой кожи. Клюв шиферно-чёрный, с ярко-выраженной беловатого цвета восковицей в основании.
У настоящих голубей, живущих в условиях дикой природы, туловище светло-серое, с беловатым надхвостьем и двумя отчётливыми тёмными полосами на внешней стороне крыльев. Последние две характеристики отличают сизого голубя от родственных ему клинтуха и вяхиря. Оперение городских и сельских птиц, являющихся одичавшими потомками различных пород домашних голубей, может иметь различные оттенки от почти белого и охристого до фиолетово-чёрного, но обычно более тёмное и без какого-либо определённого рисунка. Крылья широкие, заострённые. Хвост закруглённый, обычно с тёмным окончанием и белой окантовкой по краям. Ноги имеют оттенки от розового до серовато-чёрного, у некоторых птиц частично покрыты перьями.
Взрослая самка почти не отличается от самца, однако имеет менее насыщенный металлический отлив, который к тому же отсутствует на груди. Молодые птицы первые 6—8 месяцев жизни выглядят более тускло, без глянца, а их радужина коричневая либо серовато-коричневая.
В зависимости от размера, интенсивности окраски, цвета и ширины полосы на пояснице различают более десяти подвидов сизого голубя.
Голос
Вокализация — характерное для семейства в целом воркование: раскатистое, глухое урчание, особенно ярко выраженное у самцов в брачный период, когда они подзывают самок или метят территорию. Несколько иная тональность голоса, более спокойная и напоминающая скорее мурлыканье кошки, звучит во время насиживания яиц. В случае испуга голуби издают короткий крик «о-ррр». Вылупившиеся птенцы голубей способны издавать лишь слабые шипящие звуки, либо просто щёлкают клювом, повзрослевшие птенцы пищат при виде родителя и при кормлении. Помимо голосовой связи, птицы используют и другие способы общения, как, например, громкое хлопанье крыльями во время ухаживания или как сигнал опасности.
Органы чувств
Обладая прекрасным зрением, птица способна различать не только цвета радуги (подобно человеку и приматам), но также и ультрафиолетовые лучи. Благодаря этому, голуби могут использоваться в поисково-спасательных операциях — например, на рубеже 1980-х годов береговая охрана США провела успешные опыты по поиску людей в спасательных жилетах в открытом море. В начале эксперимента птиц обучали подавать сигнал, если они видели что-нибудь оранжевое, а затем помещали их на нижней палубе вертолёта и кружились над районом предполагаемого бедствия. Эксперимент показал, что в 93 % случаев голуби обнаруживали объект поиска, тогда как у спасателей этот показатель составил только 38 %. Слух позволяет улавливать звуки гораздо более низкой частоты, чем в состоянии услышать человек — например, шум ветра или отдалённые звуки грозы и, возможно поэтому, птицы иногда покидают территорию без какой-либо видимой причины. Хорошо известна способность сизых голубей всегда находить путь домой, где бы они ни находились — такое поведение издревле использовалось человеком для почтовой связи. Обладающие таким качеством птицы способны преодолевать до 1000 км в день. Некоторые орнитологи, и в частности, доктор Чарльз Уолкотт (Charles Walcott) из Корнелльского университета (США), связывают такое поведение со способностью улавливать магнитные поля и ориентироваться по солнцу. Их британские коллеги из Оксфорда провели эксперимент, прикрепив на спину птиц датчикиглобального позиционирования и проследив маршруты движения. Результат оказался неожиданным — голуби отдавали предпочтение земным ориентирам, таким как автомобильные дороги и железнодорожные пути, и только в незнакомой местности полагались на солнце. Наконец, голуби часто рассматриваются как достаточно развитые в интеллектуальном плане птицы. Например, японские специалисты из университета Кэйо выяснили, что эти птицы запоминают свои действия с задержкой до 5—7 секунд.
Движения
Сизый голубь, вследствие своего преимущественно скального образа жизни, не умеет усаживаться на ветви деревьев, хотя его синантропные потомки приспособились делать и это. По земле передвигается шагом, постоянно покачивая головой взад и вперёд. Иногда движется быстро и делая достаточно широкий шаг, что позволяет некоторым людям рассуждать о «жадности» птиц. Великолепно летает, достигая скорости до 185 км/ч. Особенно стремительным полётом обладают дикие птицы, обитающие в горах, — среди снежных скал нередко можно наблюдать стайки птиц, со свистом пикирующих вдоль склона в почти вертикальной плоскости. В регионах с засушливым климатом птицы опускаются к воде в глубокие колодцы. Городские голуби, более защищённые от хищников и обитающие вблизи от кормовой территории, как правило, менее искусны в воздухе.
Распространение

Ареал
Природный ареал сизого голубя занимает центральные и южные районы Евразии от Атлантики до долины Енисея, горного Алтая, Тянь-Шаня, восточной Индии и Мьянмы, а также Африку севернее Сенегала, Дарфура и побережья Аденского залива. В Европе поднимается до 54° с. ш., на Енисее до 55° с. ш., вблизи жилища человека обитает и много севернее, например, в Дудинке (69° с. ш.). За пределами материка распространён на островах Средиземного моря, Британских, Фарерских и Канарских островах, Лакшадвипе и Шри-Ланке. В Европейской части России изначально встречался лишь на побережье Азовского моря, по правому берегу Дона, Северном Кавказе и в Поволжье. Полудомашние популяции распространились почти по всему миру, поднимаясь в гораздо более холодные широты и концентрируясь вблизи жилья человека. На другие материки птицы были сознательно интродуцированы человеком — например, известно, что на американский континент голуби попали благодаря французам: колонисты завезли их в Новую Шотландию в 1606 году.
Местообитания
Биотопы обычно связаны со скалами, чаще всего на морском побережье, но также и в горах (в Гималаях достигая альпийского пояса — 4000 м и выше).[15] Предпочтение отдаёт открытым пространствам, в том числе степным и пустынным регионам (в последних селится в оазисах). Синантропные популяции селятся в нишах и в углублениях каменных и деревянных построек, занимая в первую очередь малодоступные или редко посещаемые человеком места. Как правило, оседлый вид, хотя в высокогорных районах птицы совершают незначительные вертикальные кочёвки, в холодное время года опускаясь в долины. Численность диких птиц относительно небольшая и имеет тенденцию к понижению, хотя, по мнению специалистов, она пока не достигла сколько-нибудь угрожающих размеров.
Заключения и выводы.
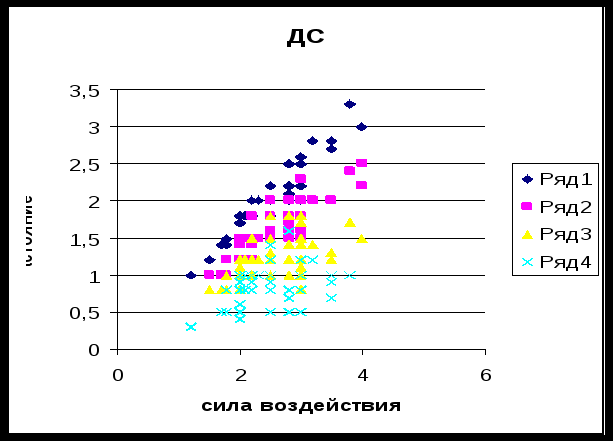
Выводы:
Чем сильнее воздействие на птиц, тем меньше ДС.
В областях с пониженным антропогенным фактором ДС больше, чем в областях где антропогенный фактор выше.
Литература:
1. Авилова К.В., Корбут В.В., Фокин С.Ю. 1994. Урбанизированная популяция водоплавающих (Anas platyrhynchos) г. Москвы. М.: 1-175
2. Вахрушев А.А., Зюзин А.А.1984. Дистанция вспугивания серой вороны в городе // Экология, биоценотическое и хозяйственное значение врановых птиц. М.: 40-42
3. Владышевский Д.В. 1975. Птицы в антропогенном ландшафте. Новосибирск:1- 199
4. Зорина З.А. 1984. Рассудочная деятельность и пластичность поведения врановых в условиях антропогенных воздействий // Экология, биоценотическое и хозяйственное значение врановых птиц. М.: 12-15
5. Ильичев В.Д., Бутьев В.Т., Константинов В.М. 1987. Птицы Москвы и Подмосковья. М.: 1-272
6. Клауснитцер Б. 1990. Экология городской среды. М.: 1-248
7. Константинов В.М., Бабенко В.Г., Барышева И.К. 1982. Численность и некоторые черты экологии синантропных популяций врановых птиц в условиях интенсивной урбанизации // Зоол.ж., 61, вып. 12: 1837-1845
8. Крушинский Л.В. 1958. Экстраполяционные рефлексы у птиц // Орнитология. М.: 145-159
9. Лысенков Е.В., Лапшин А.С., Пятаева Е.Н. 2000. Автодороги и население птиц // Мордовский орнитол.вестник, 2. Саранск: 61-68
10. Мешкова Н.Н., Федорович Е.Ю. 1996. Ориентировочно-исследовательская деятельность, подражание и игра как психологические механизмы адаптации высших позвоночных к урбанизированной среде. М.: 1-226
11. Нанкинов Д.Н., Тодоров Н.М. 1983. Исследование гибели птиц на автомобильных дорогах // Экология, № 5: 62-68
12. Рахилин В.К. 1996. Животные в жизни людей (история изучения, использования и охраны животного мира). М.: 1-264
13. Резанов А.А. 2002. К методике оценки дистанции вспугивания у птиц // Экология врановых птиц в антропогенных ландшафтах. Саранск: 100-102
14. Резанов А.А. 2003. Зона и дистанция реагирования серой вороны (Corvus cornix) на человека, как показатель уровня её толерантности к фактору беспокойства // Актуальные вопросы биологии, химии и экологии: наука и образование. М: 140-152
15. Резанов А.А. 2005. Оценка антропотолерантности птиц в условиях селитебного ландшафта // Естественнонаучное образование: методология, теория и методика. С.-Пб.: 166-170
16. Резанов А.А. 2005. Реакция серой вороны (Corvus cornix), галки (Corvus monedula) и других видов птиц на приближающейся автотранспорт // Экология врановых птиц в условиях естественных и антропогенных ландшафтов России. Казань: 140-142
17. Резанов А.А. 2006. Антропотолерантность как один из критериев синантропизации птиц // Орнитологические исследования в Северной Евразии. Ставрополь: 431-433
18. Резанов А.А. 2007. Дистанция вспугивания различных видов птиц в условиях города // Современные проблемы гуманитарных и естественных наук. Рязань: 178-179
19. Константинов В.М., Пономарев В.А. , Воронов Л.Н., Зорина З.А., Краснобаев Д.А., Лебедев И.Г., Марголин В.А., Рахимов И.И. Резанов А.А., Резанов А.Г., Родимцев А.С., Резанов А.А. и др. 2007. Серая ворона (Corvus cornix L.) в антропогенных ландшафтах Палеарктики (проблемы синантропизации и урбанизации). М.: 1-368
20. Резанов А.Г. 1998. Эволюция антропогенных кормовых методов птиц // Влияние антропогенных факторов на структуру и функционирование биоценозов и их отдельные компоненты. М.: 5-17
21. Резанов А.Г., Резанов А.А. 2002. Оценка дистанции вспугивания серой вороны (Corvus cornix) в местах с различным уровнем фактора беспокойства // Экология врановых птиц в антропогенных ландшафтах. Саранск: 102-104
22. Родимцев А.С., Шкарин В.С., Лимонова М.А., Меркулова Е.А. 1984. Зимовка врановых в г. Новокузнецке // Природа и экономика Кузбасса. Новокузнецк: 115-118
23. Телегин В.И., Ивлева Н.Г. 1983. Птицы на дорогах // Птицы Сибири. Горно-Алтайск: 256-257
24. Телегин В.И. Ивлева Н.Г. 1992. Птицы на автодорогах // Сиб.биол.ж., № 1: 53-55
25. Guillory H.D, Le Blanc D.J. 1975. Carrion feeding by birds in southwestern Louisiana // Wilson Bull. 87, N 2: 287-288
26. Kramer D.L., Bonenfant M. 1997. Direction of predator approach and the decision to flee to a refuge // Anim. Behav. 54: 289-295
27. Metcalf B.M., Davies S.J.J.F., Ladd P.G. 2000. Adaptation of behaviour by two bird species as a result of habituation to humans // Australian Bird Watcher 18, N8: 306-312
28. Protor C.J., Broom M., Ruxton C.D. 2001. Modelling antipredator vigilance and flight response in group foragers when signals are ambiguous // J. Their.Biol., 211, N 4: 409-417

Нравится материал? Поддержи автора!
Ещё документы из категории биология:
Чтобы скачать документ, порекомендуйте, пожалуйста, его своим друзьям в любой соц. сети.
После чего кнопка «СКАЧАТЬ» станет доступной!
Кнопочки находятся чуть ниже. Спасибо!
Кнопки:
Скачать документ